

Stress granules regulate stress-induced paraspeckle assembly
- PMID: 31636118
- PMCID: PMC6891081
- DOI: 10.1083/jcb.201904098
Stress granules regulate stress-induced paraspeckle assembly
Abstract
Eukaryotic cells contain a variety of RNA-protein macrocomplexes termed RNP granules. Different types of granules share multiple protein components; however, the crosstalk between spatially separated granules remains unaddressed. Paraspeckles and stress granules (SGs) are prototypical RNP granules localized exclusively in the nucleus and cytoplasm, respectively. Both granules are implicated in human diseases, such as amyotrophic lateral sclerosis. We characterized the composition of affinity-purified paraspeckle-like structures and found a significant overlap between the proteomes of paraspeckles and SGs. We further show that paraspeckle hyperassembly is typical for cells subjected to SG-inducing stresses. Using chemical and genetic disruption of SGs, we demonstrate that formation of microscopically visible SGs is required to trigger and maintain stress-induced paraspeckle assembly. Mechanistically, SGs may sequester negative regulators of paraspeckle formation, such as UBAP2L, alleviating their inhibitory effect on paraspeckles. Our study reveals a novel function for SGs as positive regulators of nuclear RNP granule assembly and suggests a role for disturbed SG-paraspeckle crosstalk in human disease.
© 2019 An et al.
Figures
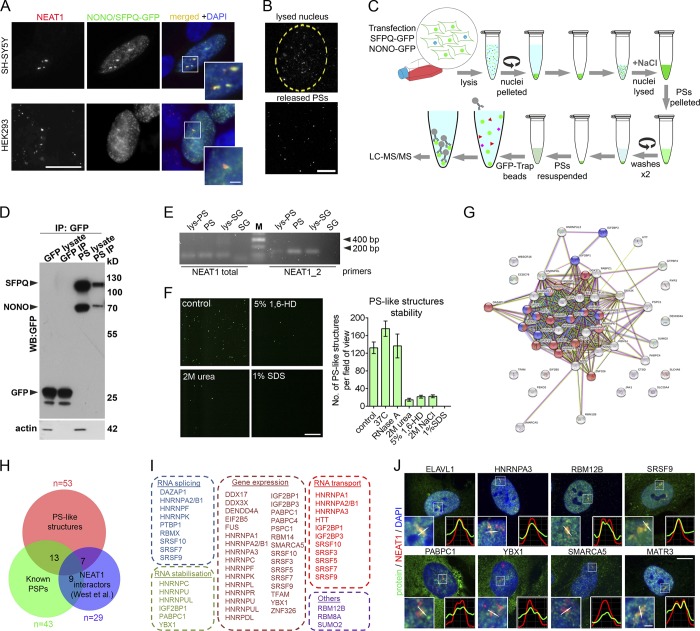
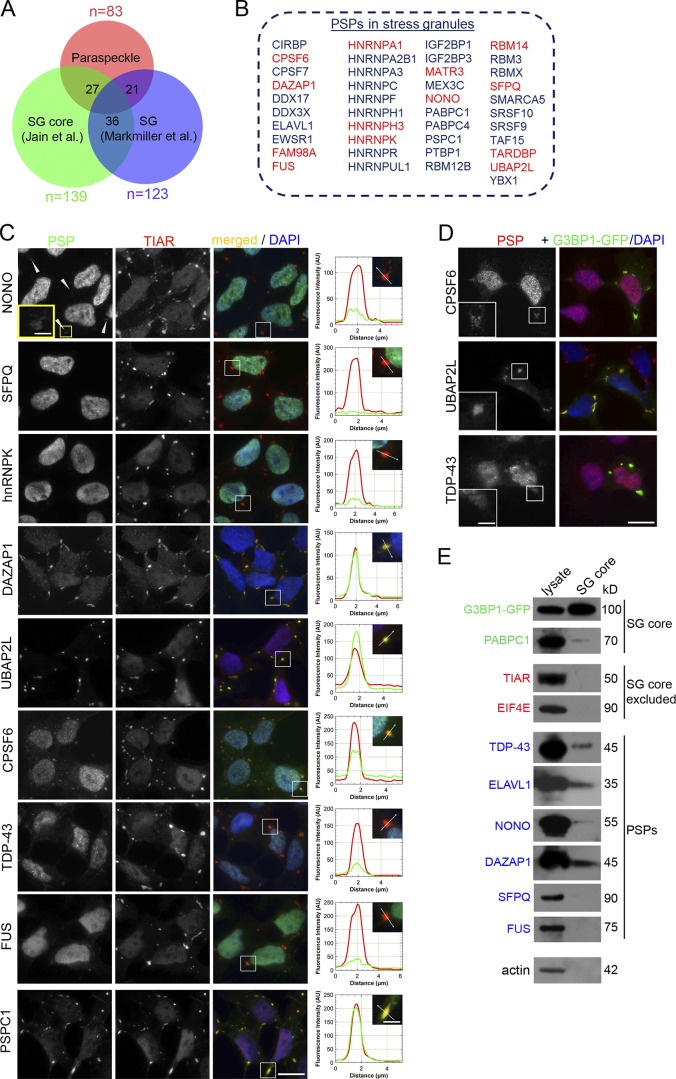
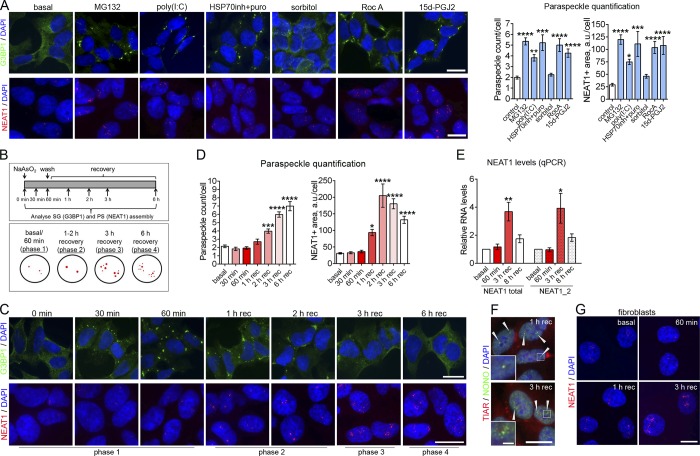
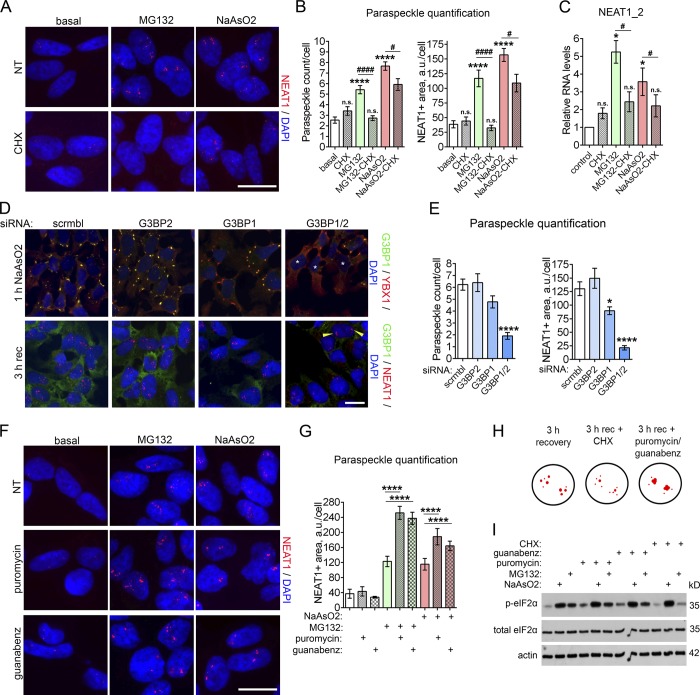

Comment in
-
Stress granules regulate paraspeckles: RNP granule continuum at work.Cell Stress. 2019 Nov 21;3(12):385-387. doi: 10.15698/cst2019.12.207. Cell Stress. 2019. PMID: 31832603 Free PMC article.
References
-
- Adriaens C., Standaert L., Barra J., Latil M., Verfaillie A., Kalev P., Boeckx B., Wijnhoven P.W., Radaelli E., Vermi W., et al. . 2016. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat. Med. 22:861–868. 10.1038/nm.4135 - DOI - PubMed
-
- An H., Skelt L., Notaro A., Highley J.R., Fox A.H., La Bella V., Buchman V.L., and Shelkovnikova T.A.. 2019. ALS-linked FUS mutations confer loss and gain of function in the nucleus by promoting excessive formation of dysfunctional paraspeckles. Acta Neuropathol. Commun. 7:7 10.1186/s40478-019-0658-x - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
