

Principles and applications of optogenetics in developmental biology
- PMID: 31641044
- PMCID: PMC6914371
- DOI: 10.1242/dev.175067
Principles and applications of optogenetics in developmental biology
Abstract
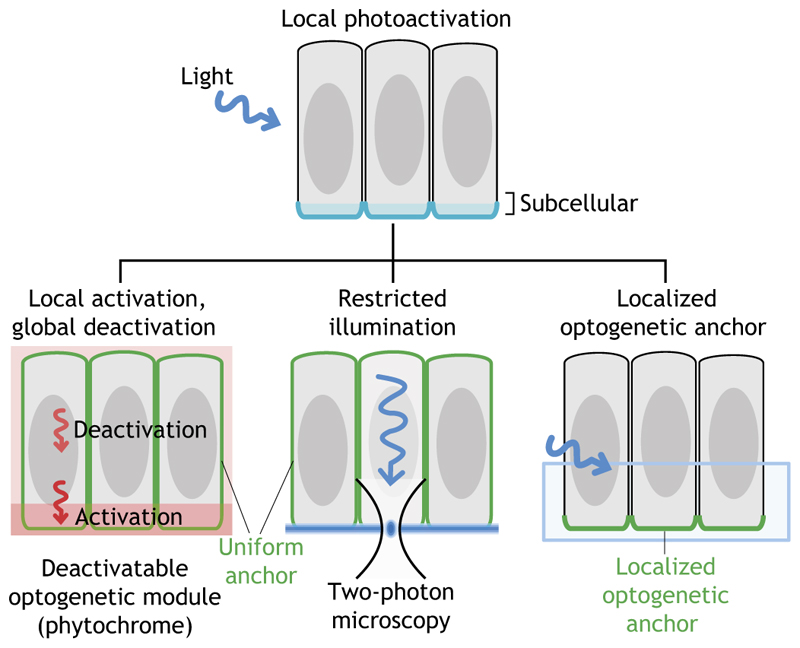
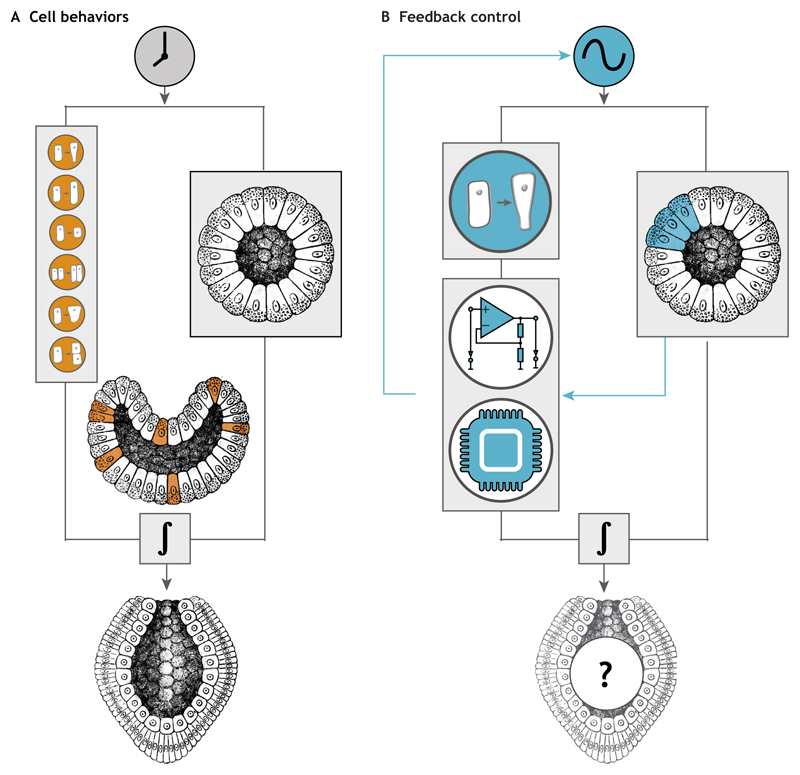
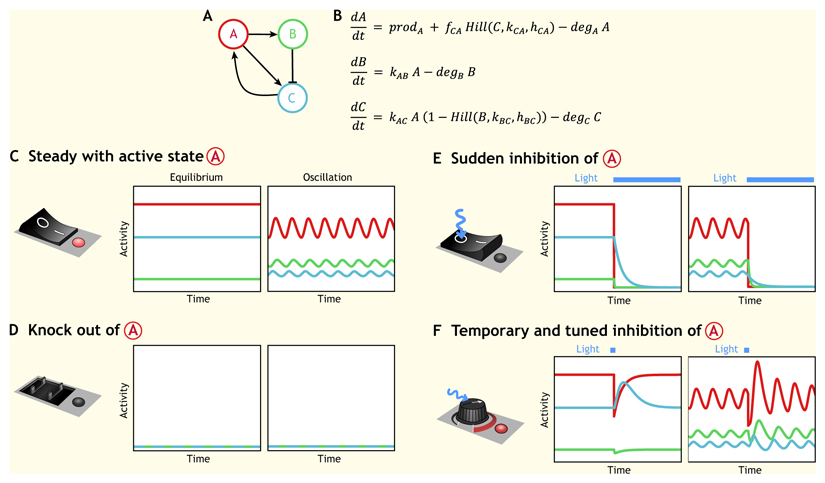
The development of multicellular organisms is controlled by highly dynamic molecular and cellular processes organized in spatially restricted patterns. Recent advances in optogenetics are allowing protein function to be controlled with the precision of a pulse of laser light in vivo, providing a powerful new tool to perturb developmental processes at a wide range of spatiotemporal scales. In this Primer, we describe the most commonly used optogenetic tools, their application in developmental biology and in the nascent field of synthetic morphogenesis.
Keywords: Embryonic development; Optogenetics; Signaling; Synthetic biology; Tissue morphogenesis.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
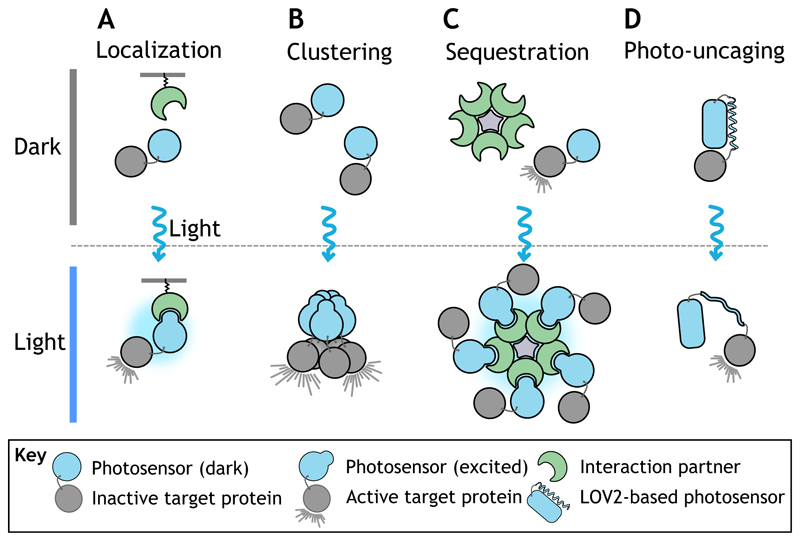
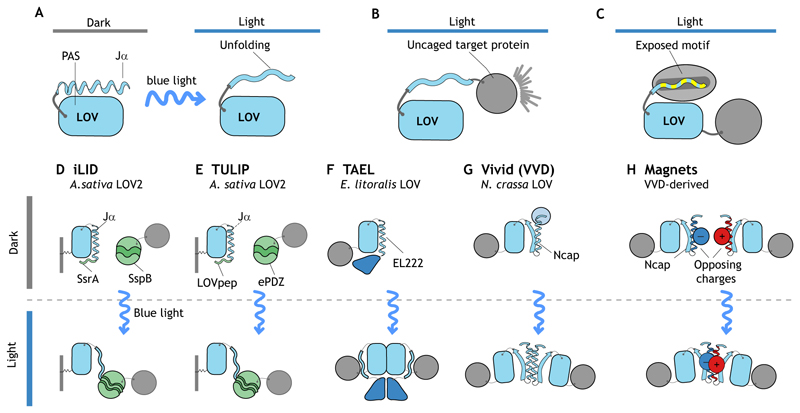
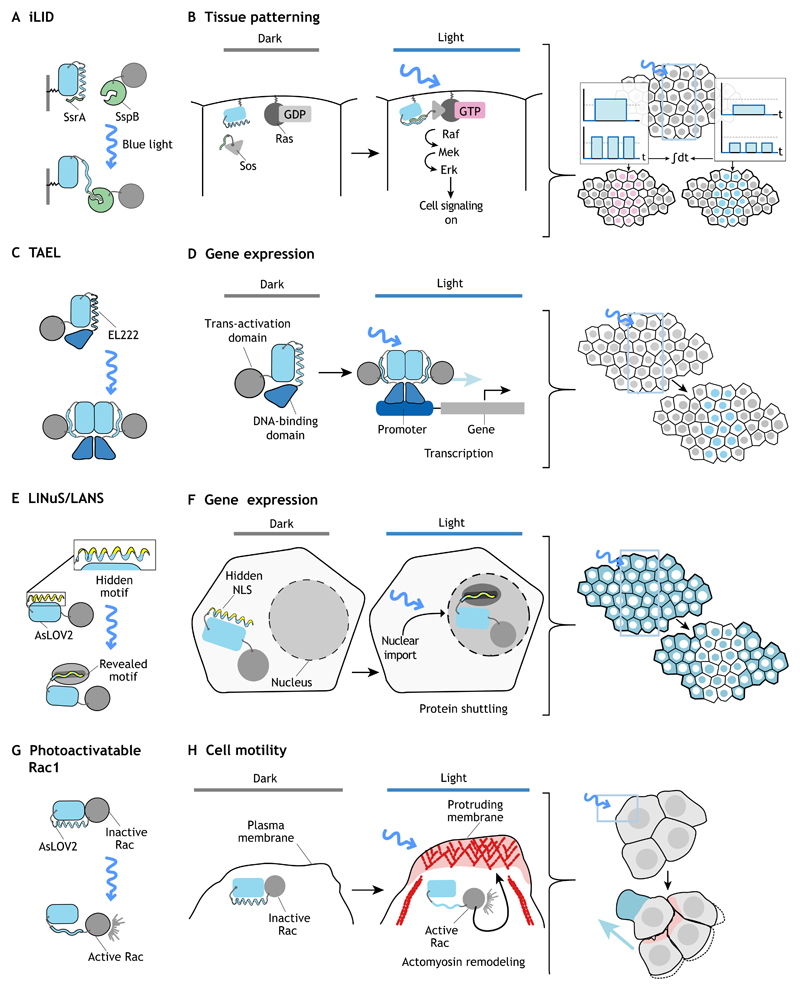
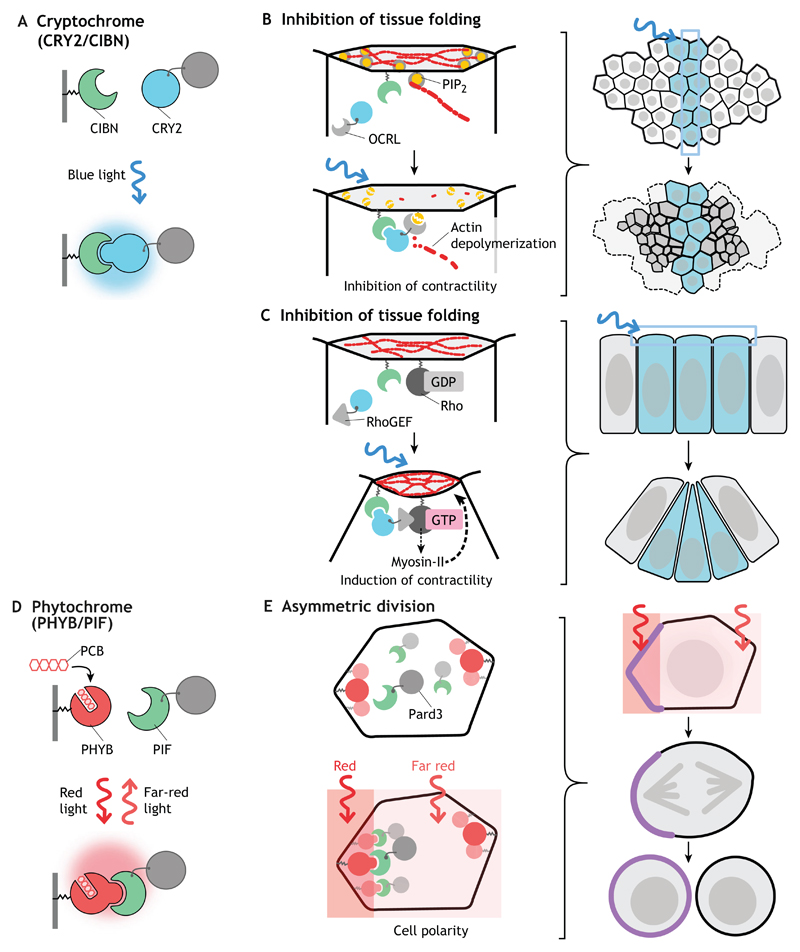
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
