

Protochromic absorption changes in the two-cysteine photocycle of a blue/orange cyanobacteriochrome
- PMID: 31649035
- PMCID: PMC6901293
- DOI: 10.1074/jbc.RA119.010384
Protochromic absorption changes in the two-cysteine photocycle of a blue/orange cyanobacteriochrome
Erratum in
-
Correction: Protochromic absorption changes in the two-cysteine photocycle of a blue/orange cyanobacteriochrome.J Biol Chem. 2020 Feb 7;295(6):1767. doi: 10.1074/jbc.AAC120.012567. J Biol Chem. 2020. PMID: 32034012 Free PMC article. No abstract available.
Abstract
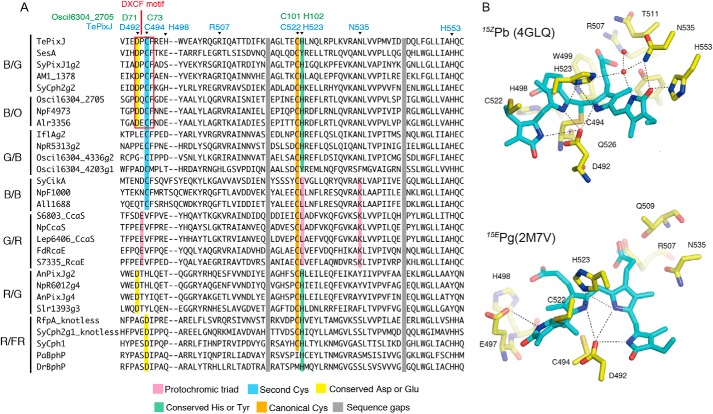
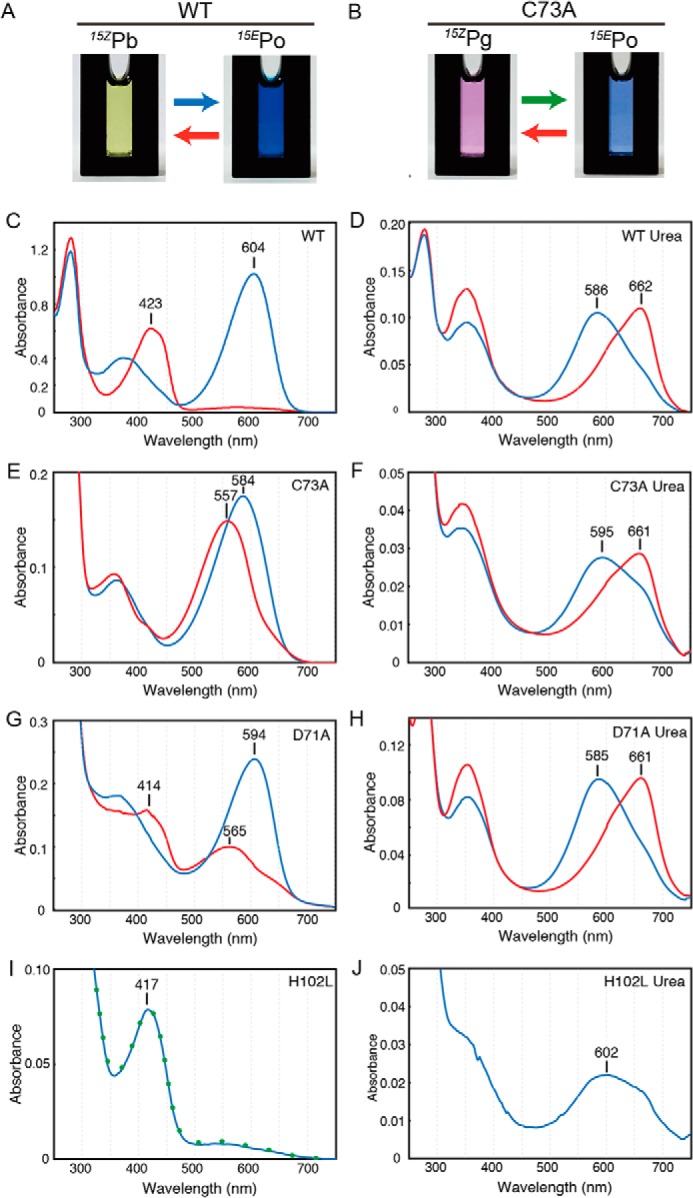
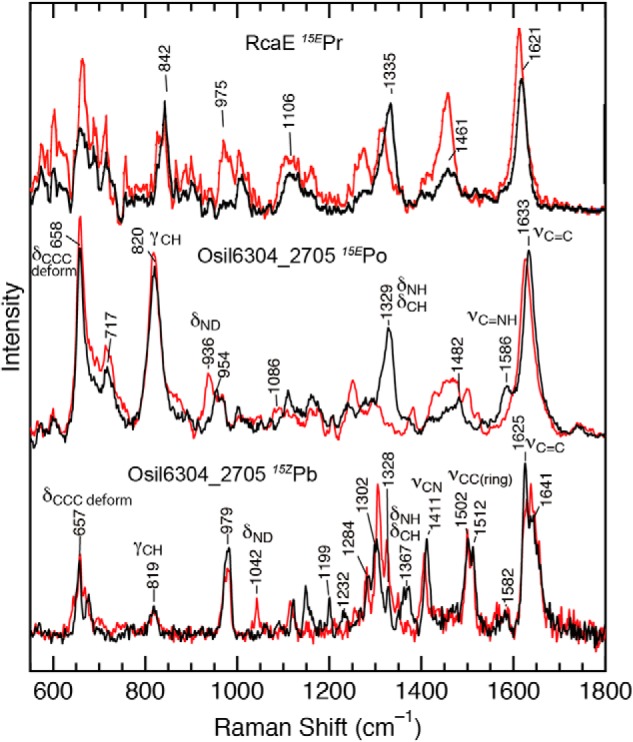
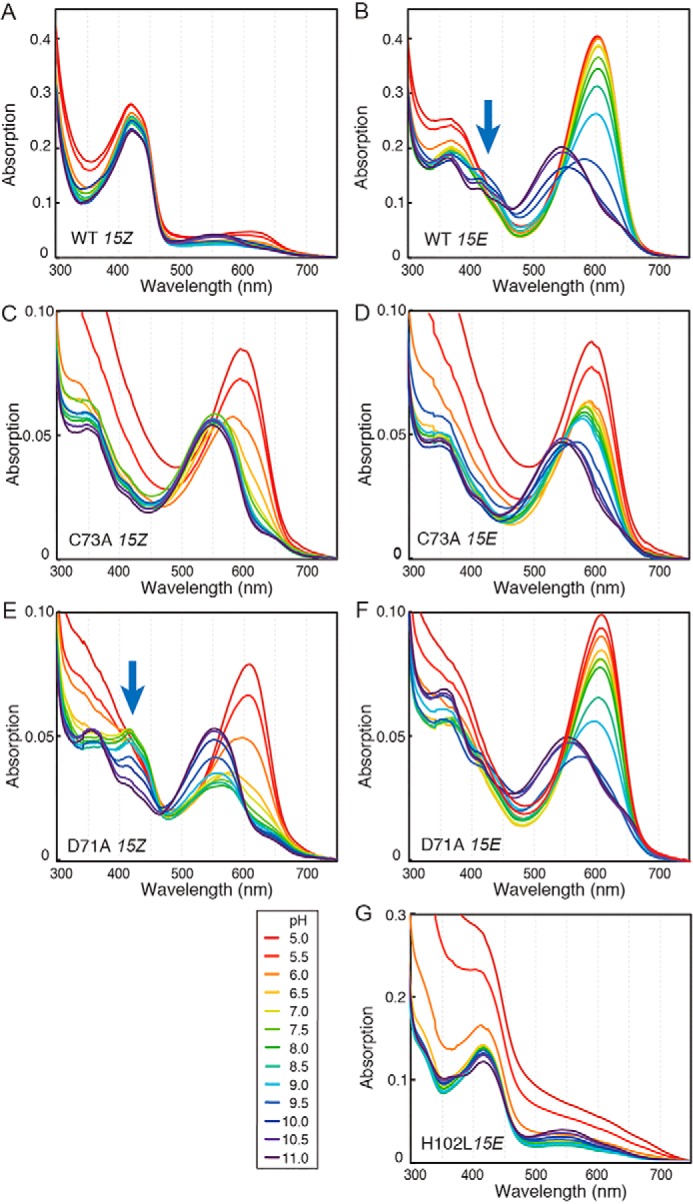
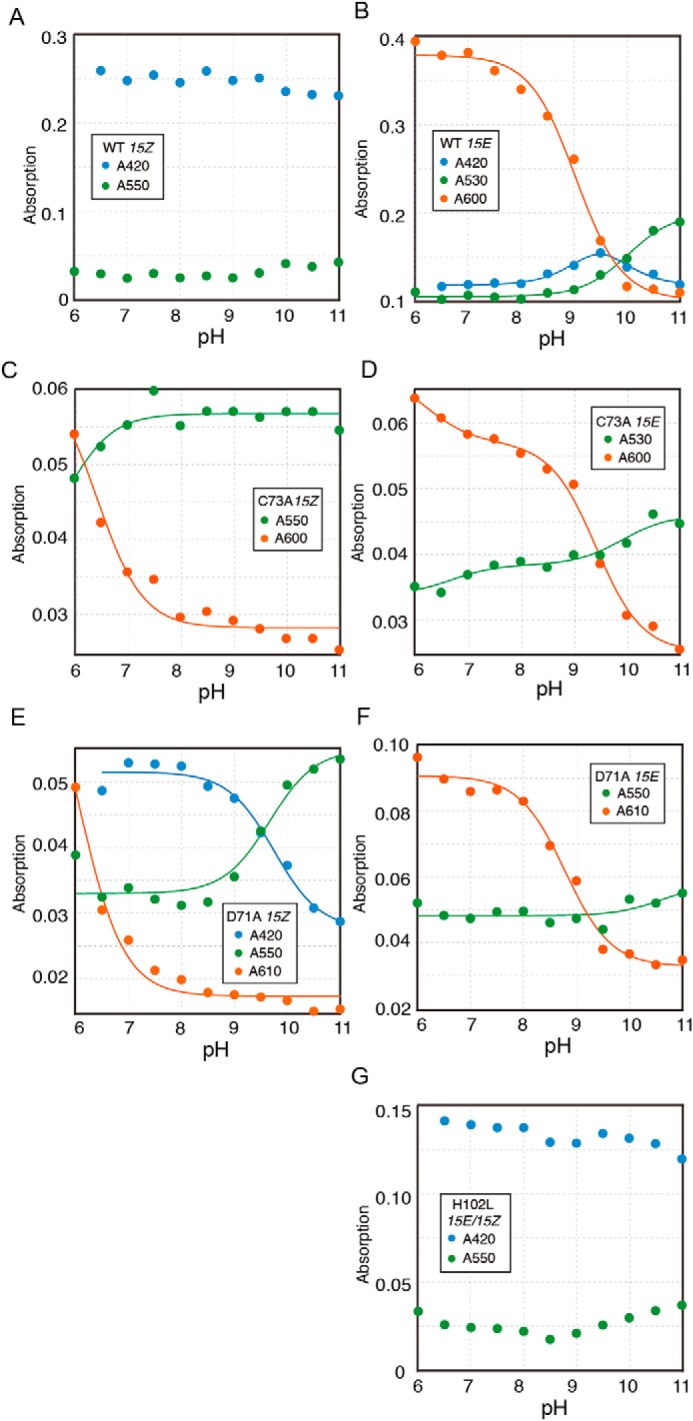
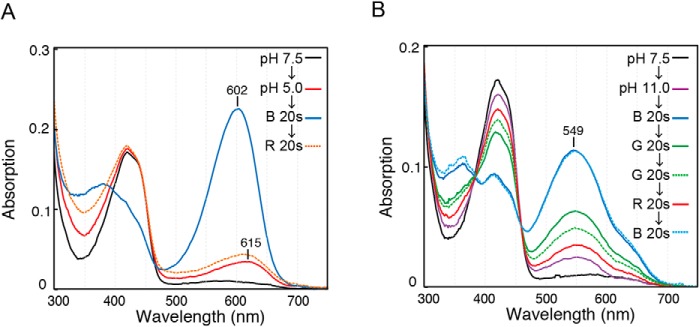
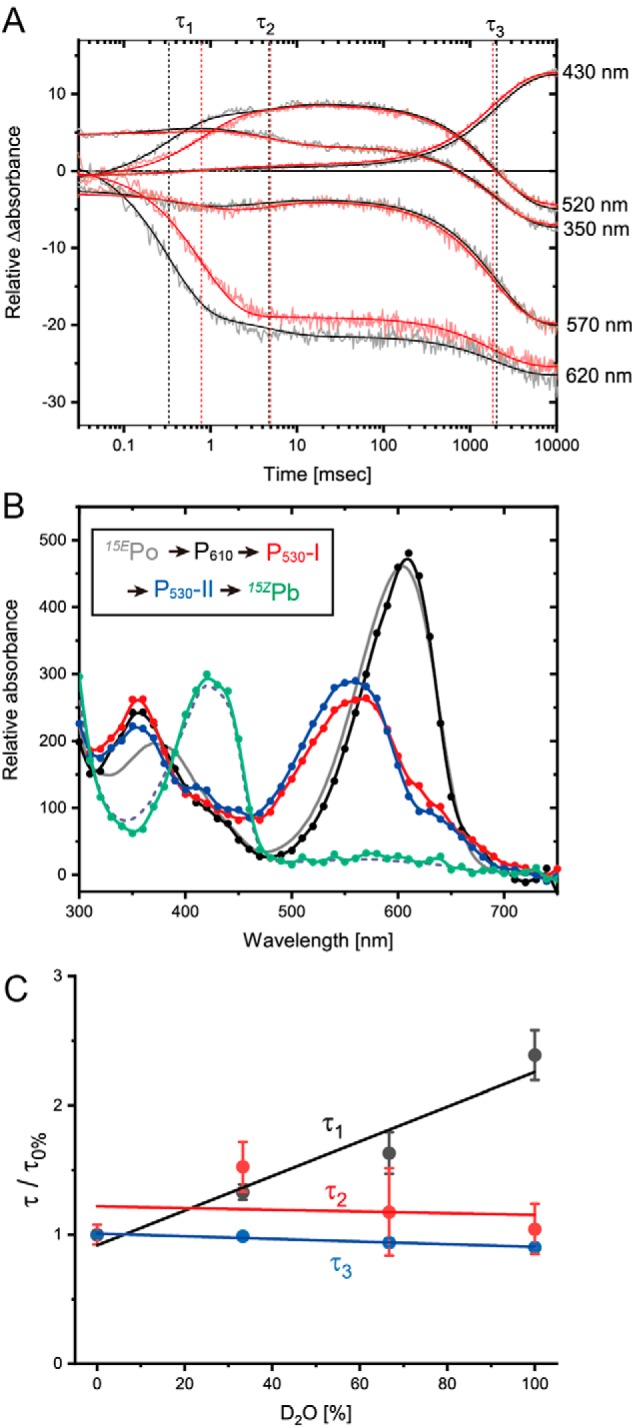
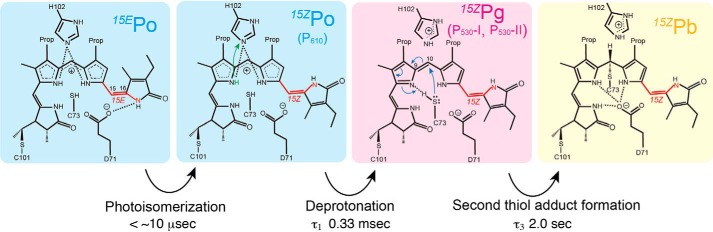
Cyanobacteriochromes (CBCRs) are phytochrome-related photosensors with diverse spectral sensitivities spanning the entire visible spectrum. They covalently bind bilin chromophores via conserved cysteine residues and undergo 15Z/15E bilin photoisomerization upon light illumination. CBCR subfamilies absorbing violet-blue light use an additional cysteine residue to form a second bilin-thiol adduct in a two-Cys photocycle. However, the process of second thiol adduct formation is incompletely understood, especially the involvement of the bilin protonation state. Here, we focused on the Oscil6304_2705 protein from the cyanobacterium Oscillatoria acuminata PCC 6304, which photoconverts between a blue-absorbing 15Z state ( 15Z Pb) and orange-absorbing 15E state ( 15E Po). pH titration analysis revealed that 15Z Pb was stable over a wide pH range, suggesting that bilin protonation is stabilized by a second thiol adduct. As revealed by resonance Raman spectroscopy, 15E Po harbored protonated bilin at both acidic and neutral pH, but readily converted to a deprotonated green-absorbing 15Z state ( 15Z Pg) at alkaline pH. Site-directed mutagenesis revealed that the conserved Asp-71 and His-102 residues are required for second thiol adduct formation in 15Z Pb and bilin protonation in 15E Po, respectively. An Oscil6304_2705 variant lacking the second cysteine residue, Cys-73, photoconverted between deprotonated 15Z Pg and protonated 15E Pr, similarly to the protochromic photocycle of the green/red CBCR subfamily. Time-resolved spectroscopy revealed 15Z Pg formation as an intermediate in the 15E Pr-to- 15Z Pg conversion with a significant solvent-isotope effect, suggesting the sequential occurrence of 15EP-to-15Z photoisomerization, deprotonation, and second thiol adduct formation. Our findings uncover the details of protochromic absorption changes underlying the two-Cys photocycle of violet-blue-absorbing CBCR subfamilies.
Keywords: Raman spectroscopy; bilin; cyanobacteria; cyanobacteriochrome; photobiology; photoreceptor; phytochrome; spectroscopy; time-resolved spectroscopy.
© 2019 Sato et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
