

MAIT cells are imprinted by the microbiota in early life and promote tissue repair
- PMID: 31649166
- PMCID: PMC7603427
- DOI: 10.1126/science.aax6624
MAIT cells are imprinted by the microbiota in early life and promote tissue repair
Abstract
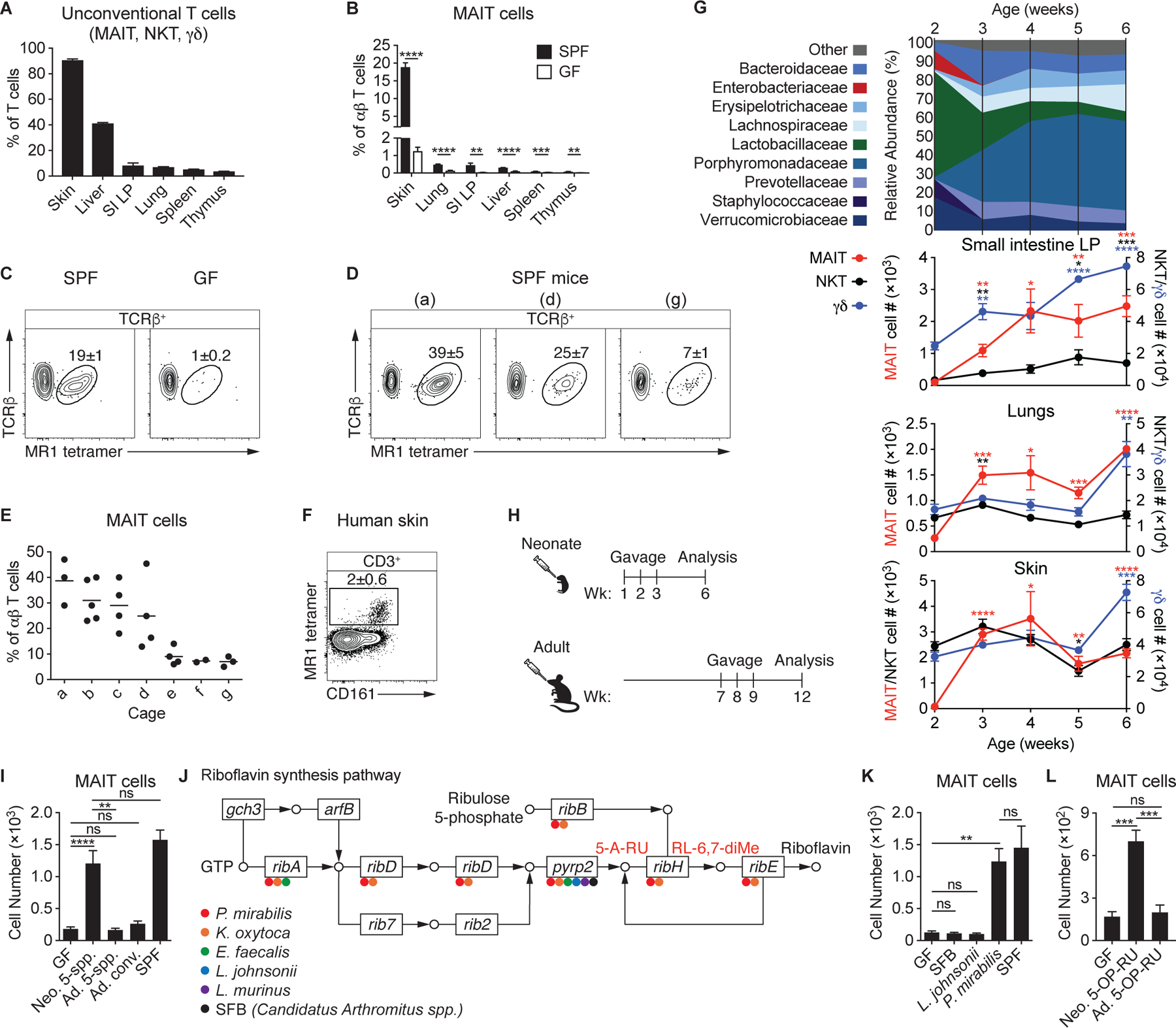
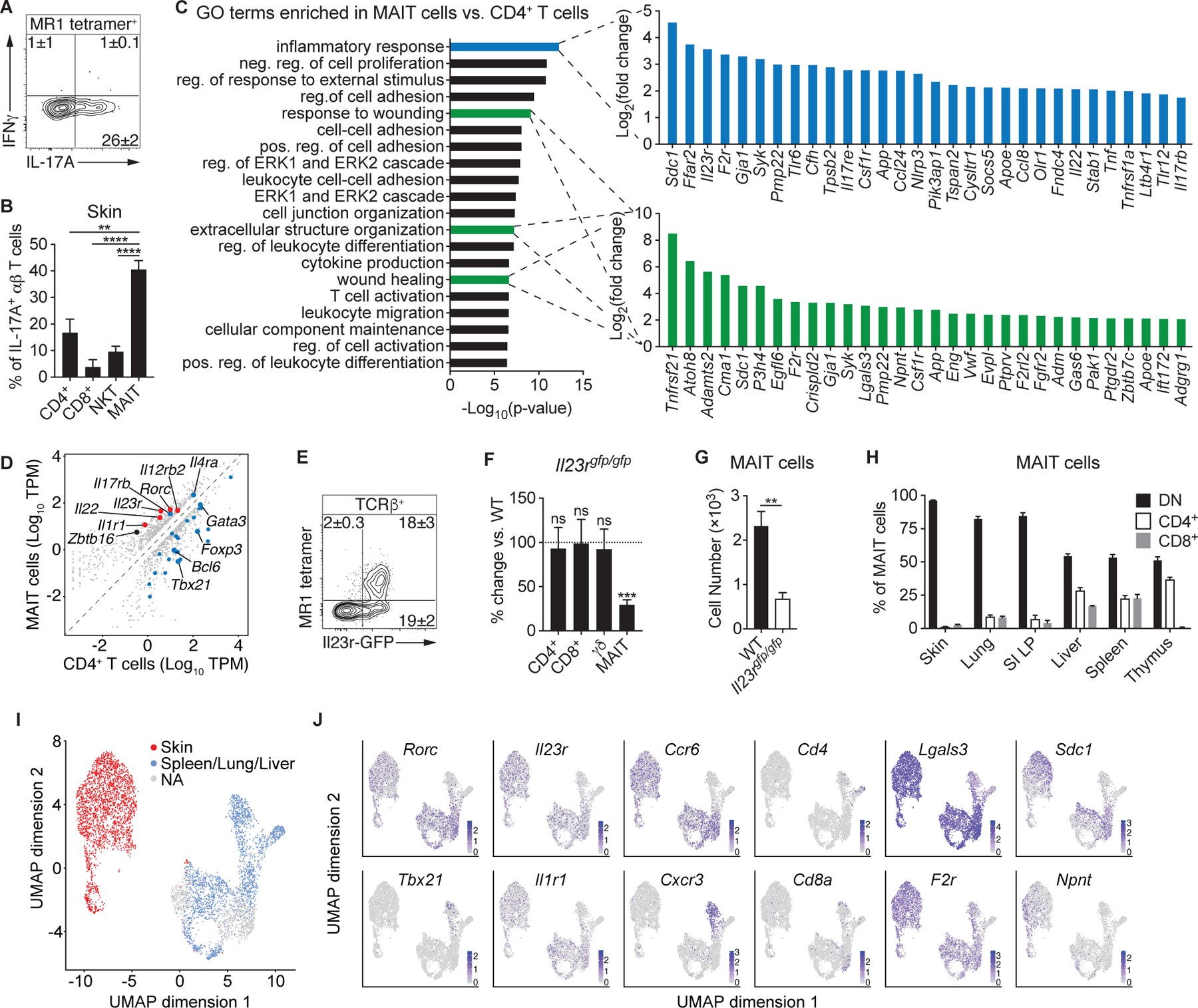
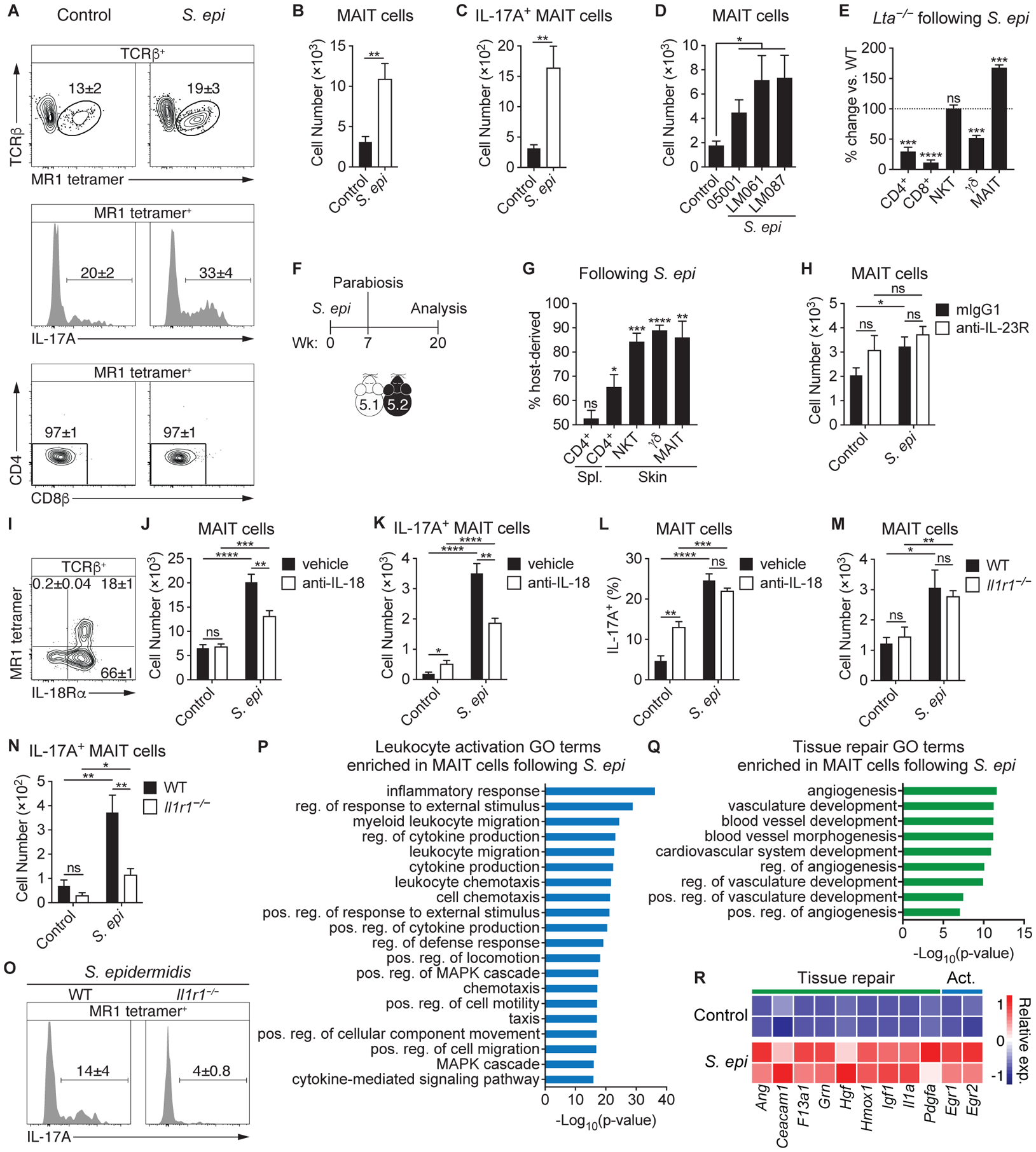
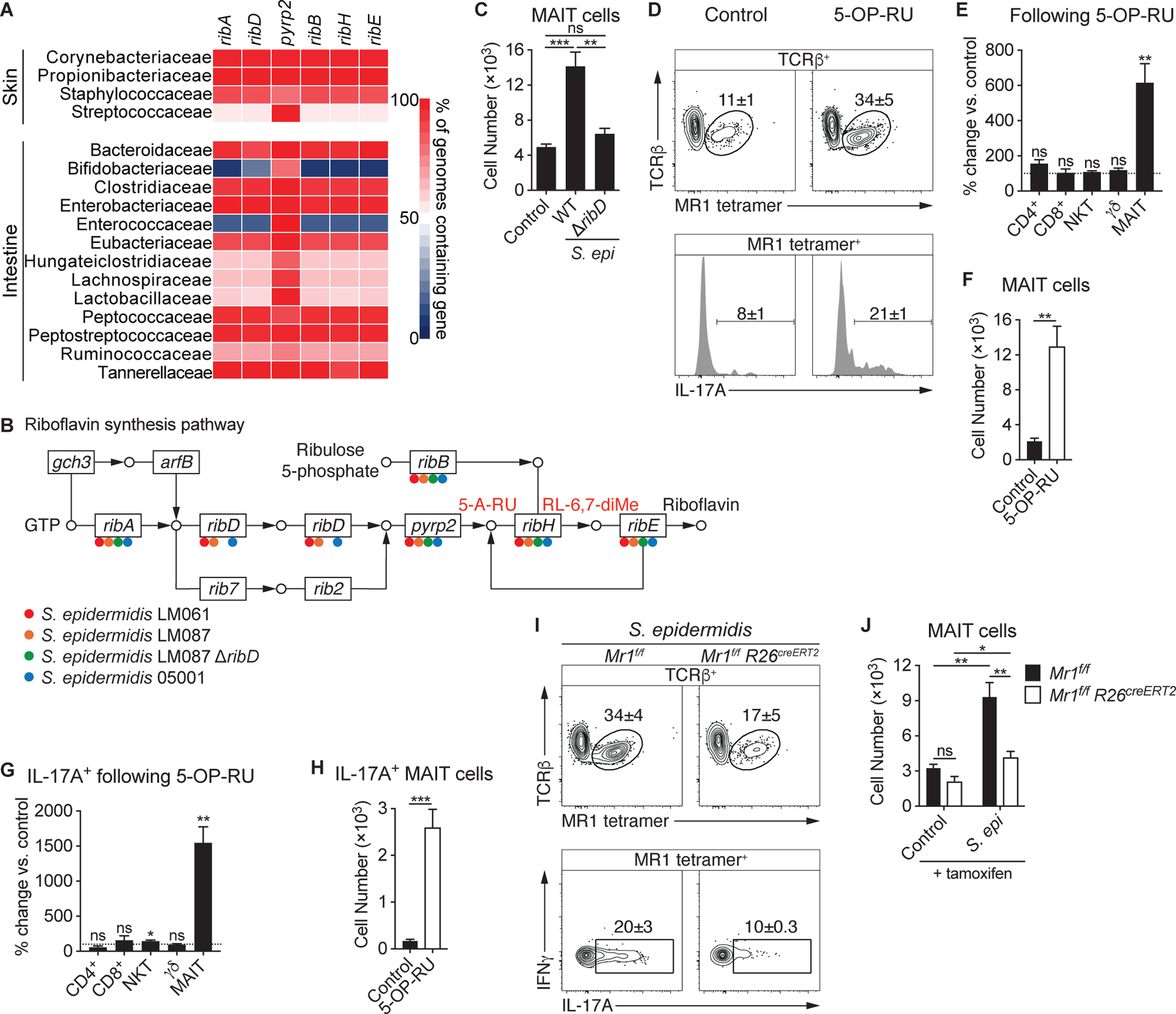
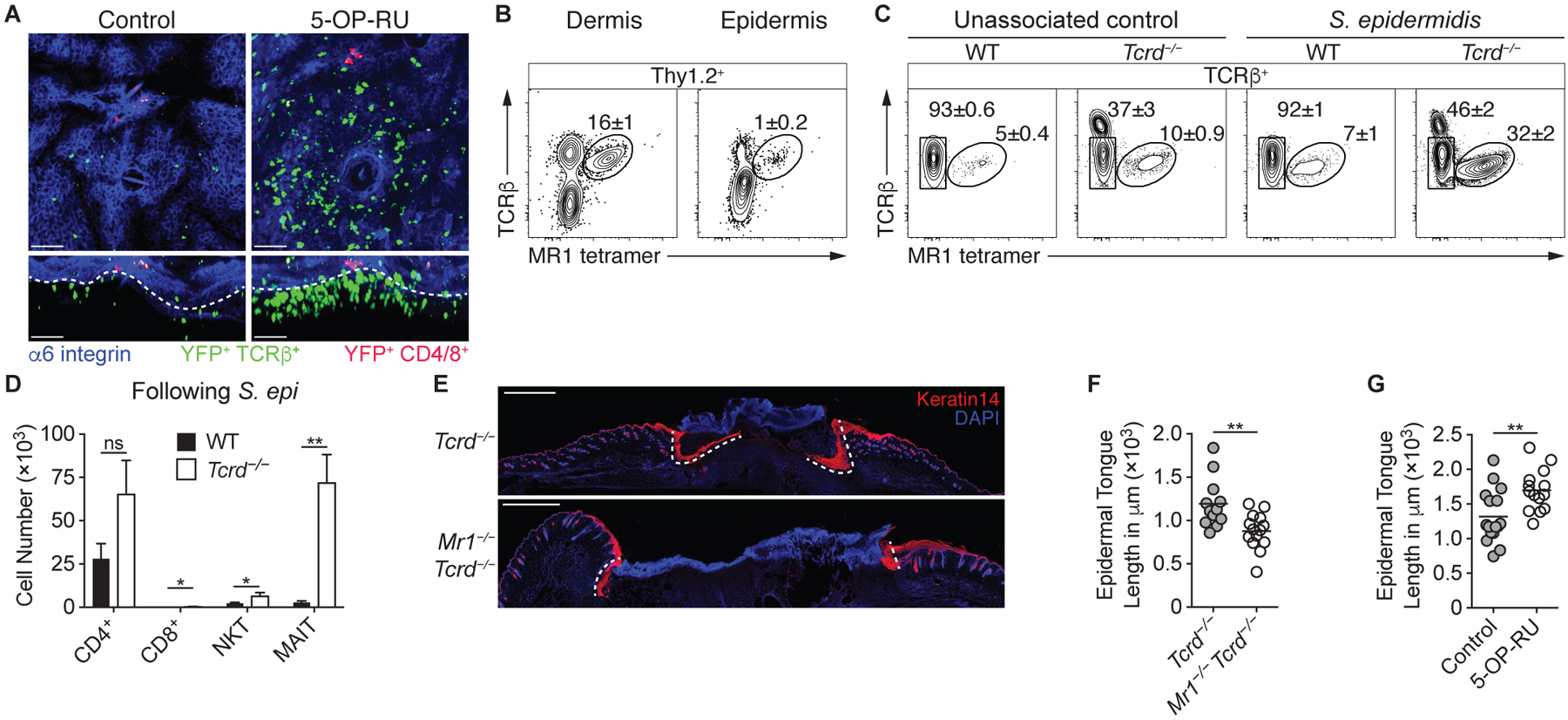
How early-life colonization and subsequent exposure to the microbiota affect long-term tissue immunity remains poorly understood. Here, we show that the development of mucosal-associated invariant T (MAIT) cells relies on a specific temporal window, after which MAIT cell development is permanently impaired. This imprinting depends on early-life exposure to defined microbes that synthesize riboflavin-derived antigens. In adults, cutaneous MAIT cells are a dominant population of interleukin-17A (IL-17A)-producing lymphocytes, which display a distinct transcriptional signature and can subsequently respond to skin commensals in an IL-1-, IL-18-, and antigen-dependent manner. Consequently, local activation of cutaneous MAIT cells promotes wound healing. Together, our work uncovers a privileged interaction between defined members of the microbiota and MAIT cells, which sequentially controls both tissue-imprinting and subsequent responses to injury.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Immune cells for microbiota surveillance.Science. 2019 Oct 25;366(6464):419-420. doi: 10.1126/science.aaz4014. Science. 2019. PMID: 31649181 No abstract available.
Similar articles
-
Microbial metabolites control the thymic development of mucosal-associated invariant T cells.Science. 2019 Oct 25;366(6464):494-499. doi: 10.1126/science.aaw2719. Epub 2019 Aug 29. Science. 2019. PMID: 31467190
-
MAIT cells and the microbiome.Front Immunol. 2023 Feb 23;14:1127588. doi: 10.3389/fimmu.2023.1127588. eCollection 2023. Front Immunol. 2023. PMID: 36911683 Free PMC article. Review.
-
Functional Heterogeneity and Antimycobacterial Effects of Mouse Mucosal-Associated Invariant T Cells Specific for Riboflavin Metabolites.J Immunol. 2015 Jul 15;195(2):587-601. doi: 10.4049/jimmunol.1402545. Epub 2015 Jun 10. J Immunol. 2015. PMID: 26063000 Free PMC article.
-
Neuropathologic, phenotypic and functional analyses of Mucosal Associated Invariant T cells in Multiple Sclerosis.Clin Immunol. 2016 May;166-167:1-11. doi: 10.1016/j.clim.2016.03.014. Epub 2016 Apr 2. Clin Immunol. 2016. PMID: 27050759
-
MAIT cells: Conserved watchers on the wall.J Exp Med. 2025 Jan 6;222(1):e20232298. doi: 10.1084/jem.20232298. Epub 2024 Oct 24. J Exp Med. 2025. PMID: 39446132 Free PMC article. Review.
Cited by
-
Postnatal Expansion, Maturation, and Functionality of MR1T Cells in Humans.Front Immunol. 2020 Sep 16;11:556695. doi: 10.3389/fimmu.2020.556695. eCollection 2020. Front Immunol. 2020. PMID: 33042140 Free PMC article.
-
How MHCII signaling promotes benign host-microbiota interactions.PLoS Pathog. 2020 Jun 29;16(6):e1008558. doi: 10.1371/journal.ppat.1008558. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32598378 Free PMC article. Review. No abstract available.
-
PTEN directs developmental and metabolic signaling for innate-like T cell fate and tissue homeostasis.Nat Cell Biol. 2022 Nov;24(11):1642-1654. doi: 10.1038/s41556-022-01011-w. Epub 2022 Oct 27. Nat Cell Biol. 2022. PMID: 36302969 Free PMC article.
-
MR1-Restricted MAIT Cells From The Human Lung Mucosal Surface Have Distinct Phenotypic, Functional, and Transcriptomic Features That Are Preserved in HIV Infection.Front Immunol. 2021 Apr 9;12:631410. doi: 10.3389/fimmu.2021.631410. eCollection 2021. Front Immunol. 2021. PMID: 33897687 Free PMC article.
-
The γδ IEL effector API5 masks genetic susceptibility to Paneth cell death.Nature. 2022 Oct;610(7932):547-554. doi: 10.1038/s41586-022-05259-y. Epub 2022 Oct 5. Nature. 2022. PMID: 36198790 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
