

Integrating the underlying structure of stochasticity into community ecology
- PMID: 31652337
- PMCID: PMC7027466
- DOI: 10.1002/ecy.2922
Integrating the underlying structure of stochasticity into community ecology
Abstract
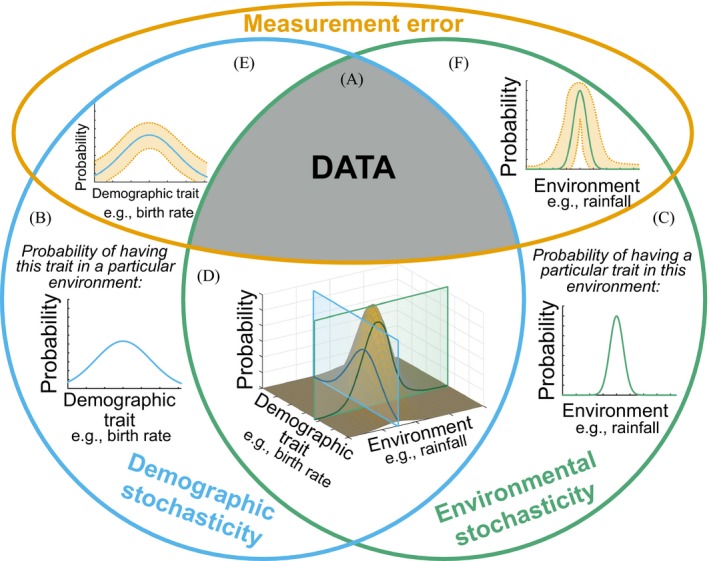
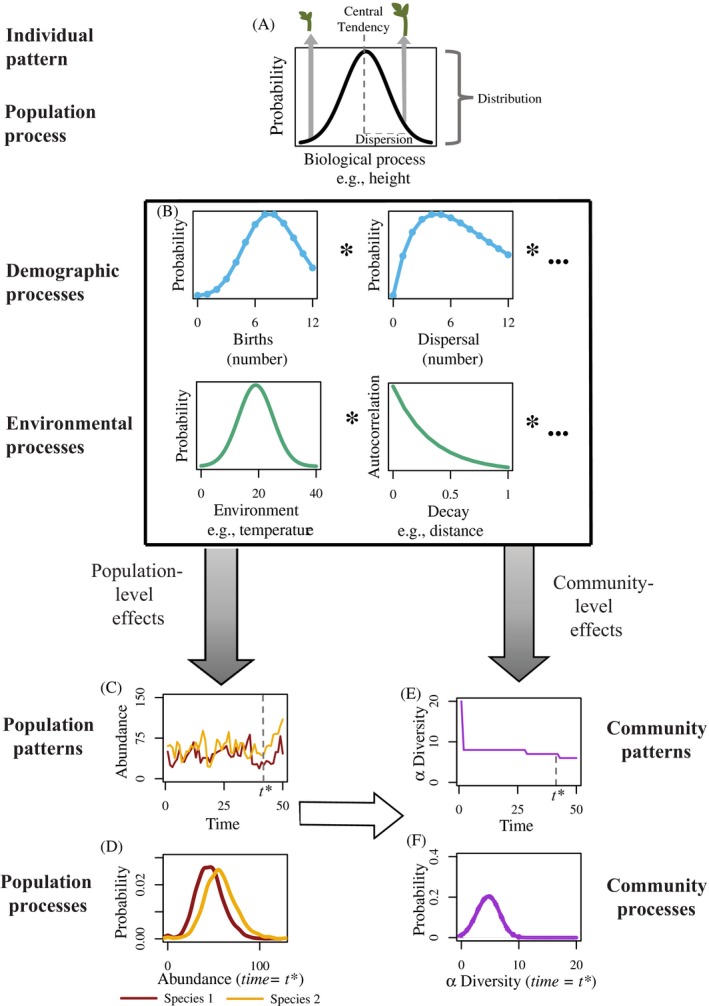
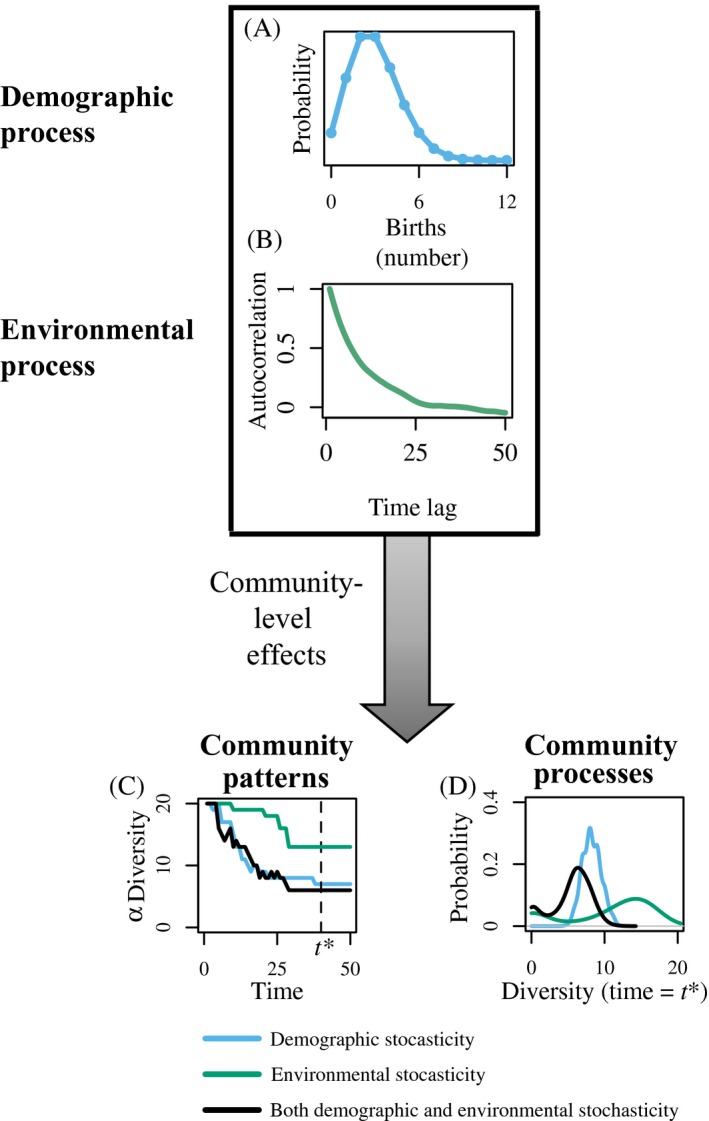
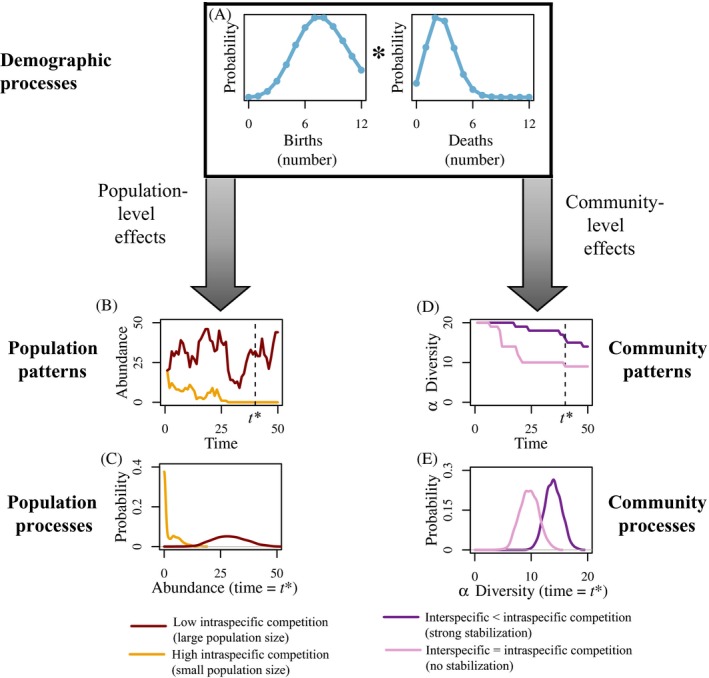
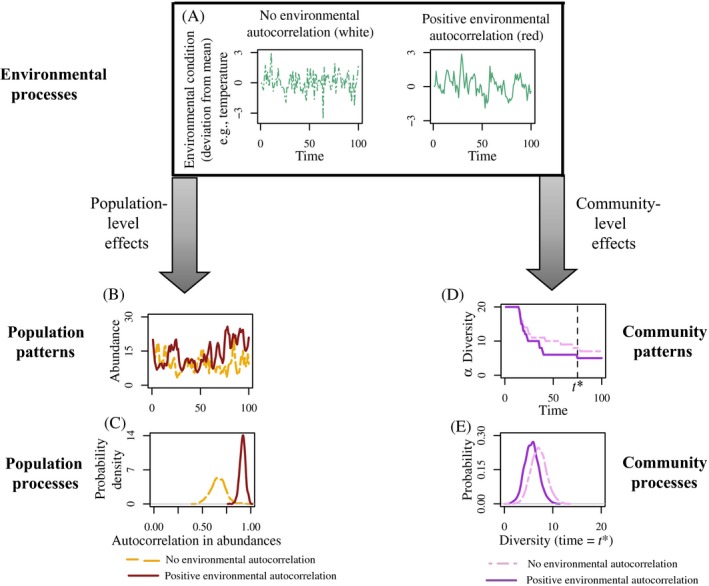
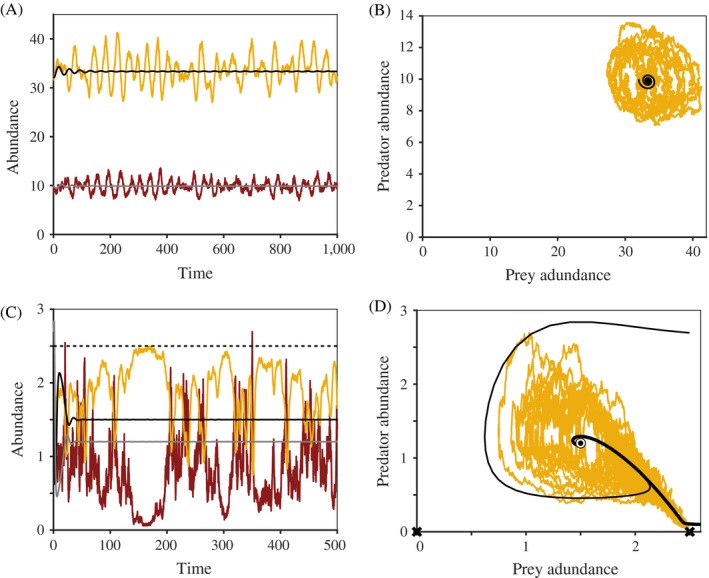
Stochasticity is a core component of ecology, as it underlies key processes that structure and create variability in nature. Despite its fundamental importance in ecological systems, the concept is often treated as synonymous with unpredictability in community ecology, and studies tend to focus on single forms of stochasticity rather than taking a more holistic view. This has led to multiple narratives for how stochasticity mediates community dynamics. Here, we present a framework that describes how different forms of stochasticity (notably demographic and environmental stochasticity) combine to provide underlying and predictable structure in diverse communities. This framework builds on the deep ecological understanding of stochastic processes acting at individual and population levels and in modules of a few interacting species. We support our framework with a mathematical model that we use to synthesize key literature, demonstrating that stochasticity is more than simple uncertainty. Rather, stochasticity has profound and predictable effects on community dynamics that are critical for understanding how diversity is maintained. We propose next steps that ecologists might use to explore the role of stochasticity for structuring communities in theoretical and empirical systems, and thereby enhance our understanding of community dynamics.
Keywords: autocorrelation; demographic stochasticity; distribution; diversity; environmental stochasticity; population dynamics; scale; uncertainty.
© 2019 The Authors. Ecology published by Wiley Periodicals, Inc. on behalf of Ecological Society of America.
Figures
References
-
- Abbott, K. C. , and Nolting B. C.. 2017. Alternative (un)stable states in a stochastic predator–prey model. Ecological Complexity 32:181–195.
-
- Adler, P. B. , and Drake J. M.. 2008. Environmental variation, stochastic extinction, and competitive coexistence. American Naturalist 172:E186–E195. - PubMed
-
- Amarasekare, P. 2006. Productivity, dispersal and the coexistence of intraguild predators and prey. Journal of Theoretical Biology 243:121–133. - PubMed
Publication types
MeSH terms
Grants and funding
- DFG FZT 118/sDiv, the Synthesis Centre of iDiv/International
- XDPB0203/Chinese Academy of Sciences/International
- 1644641/Division of Environmental Biology/International
- M.L. 2016, Chp. 186, Sec. 2, Subd. 08b/Legislative-Citizen Commission on Minnesota Resources (LCCMR)/International
- FT140100498/Australian Research Council/International
- University of Minnesota/International
- FT140100498/James S. McDonnell Foundation/International
- 31722010/James S. McDonnell Foundation/International
- XDPB0203/James S. McDonnell Foundation/International
- 31570426/James S. McDonnell Foundation/International
- 31622014/James S. McDonnell Foundation/International
- 2013/James S. McDonnell Foundation/International
- 1474/James S. McDonnell Foundation/International
- Chinese Academy of Sciences/International
- 31570426 31622014 31722010/National Natural Science Foundation of China/International
- Irish Research Council/International