

Condensin II protein dysfunction impacts mitochondrial respiration and mitochondrial oxidative stress responses
- PMID: 31653782
- PMCID: PMC6899004
- DOI: 10.1242/jcs.233783
Condensin II protein dysfunction impacts mitochondrial respiration and mitochondrial oxidative stress responses
Abstract
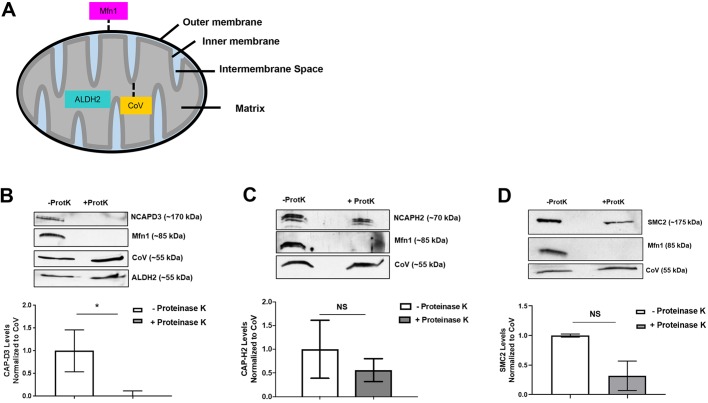
The maintenance of mitochondrial respiratory function and homeostasis is essential to human health. Here, we identify condensin II subunits as novel regulators of mitochondrial respiration and mitochondrial stress responses. Condensin II is present in the nucleus and cytoplasm. While the effects of condensin II depletion on nuclear genome organization are well studied, the effects on essential cytoplasmic and metabolic processes are not as well understood. Excitingly, we observe that condensin II chromosome-associated protein (CAP) subunits individually localize to different regions of mitochondria, suggesting possible mitochondrial-specific functions independent from those mediated by the canonical condensin II holocomplex. Changes in cellular ATP levels and mitochondrial respiration are observed in condensin II CAP subunit-deficient cells. Surprisingly, we find that loss of NCAPD3 also sensitizes cells to oxidative stress. Together, these studies identify new, and possibly independent, roles for condensin II CAP subunits in preventing mitochondrial damage and dysfunction. These findings reveal a new area of condensin protein research that could contribute to the identification of targets to treat diseases where aberrant function of condensin II proteins is implicated.
Keywords: Condensin; Mitochondria; Oxidative stress.
© 2019. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures







References
-
- Bantug G. R., Fischer M., Grählert J., Balmer M. L., Unterstab G., Develioglu L., Steiner R., Zhang L., Costa A. S. H., Gubser P. M. et al. (2018). Mitochondria-endoplasmic reticulum contact sites function as immunometabolic hubs that orchestrate the rapid recall response of memory CD8+ T cells. Immunity 48, 542-555.e6. 10.1016/j.immuni.2018.02.012 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
