

Recall Responses from Brain-Resident Memory CD8+ T Cells (bTRM) Induce Reactive Gliosis
- PMID: 31655062
- PMCID: PMC6807101
- DOI: 10.1016/j.isci.2019.10.005
Recall Responses from Brain-Resident Memory CD8+ T Cells (bTRM) Induce Reactive Gliosis
Abstract
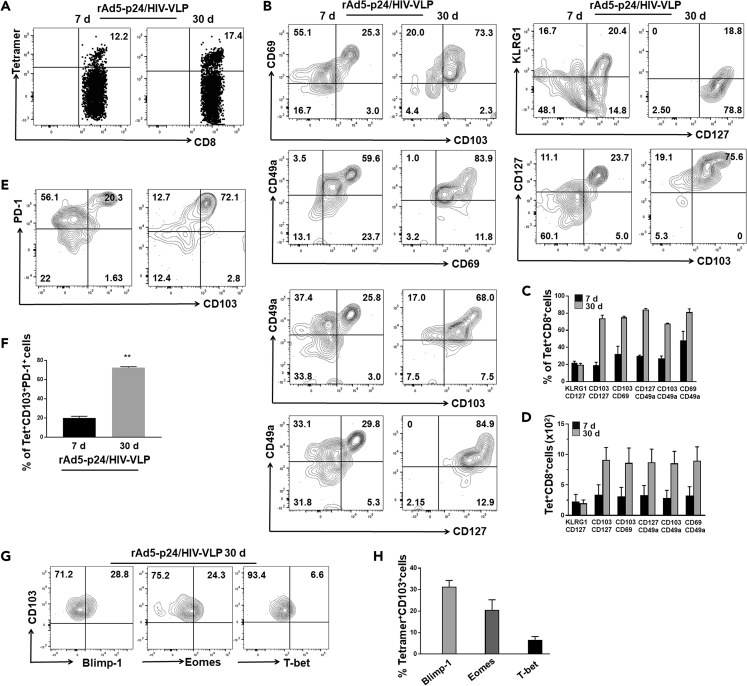
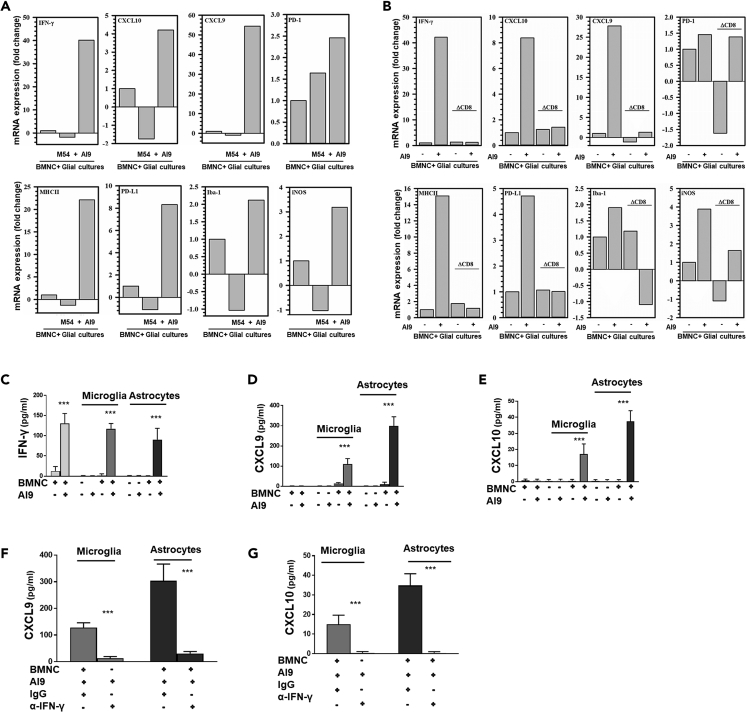
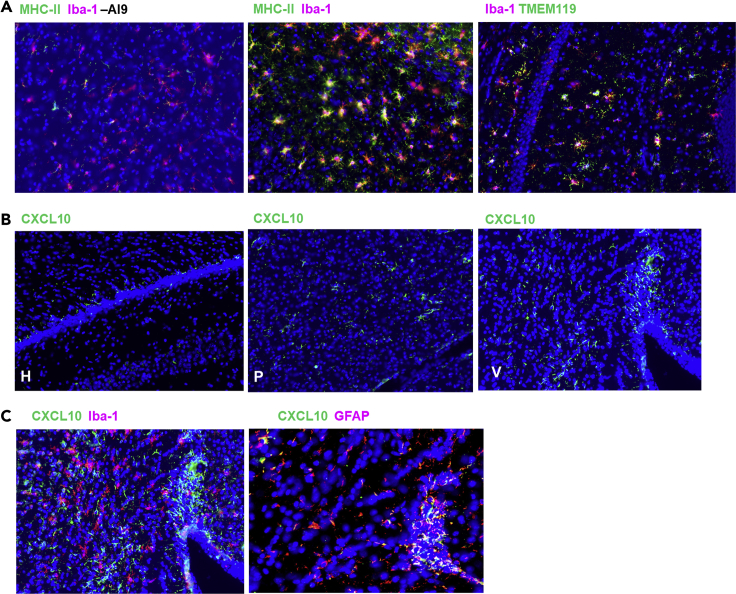
HIV-associated neurocognitive disorders (HAND) persist even during effective combination antiretroviral therapy (cART). Although the cause of HAND is unknown, studies link chronic immune activation, neuroinflammation, and cerebrospinal fluid viral escape to disease progression. In this study, we tested the hypothesis that specific, recall immune responses from brain-resident memory T cells (bTRM) could activate glia and induce neurotoxic mediators. To address this question, we developed a heterologous prime-central nervous system (CNS) boost strategy in mice. We observed that the murine brain became populated with long-lived CD8+ bTRM, some being specific for an immunodominant Gag epitope. Recall stimulation using HIV-1 AI9 peptide administered in vivo resulted in microglia displaying elevated levels of major histocompatibility complex class II and programmed death-ligand 1, and demonstrating tissue-wide reactive gliosis. Immunostaining further confirmed this glial activation. Taken together, these results indicate that specific, adaptive recall responses from bTRM can induce reactive gliosis and production of neurotoxic mediators.
Keywords: Immune System Disorder; Immunology; Neuroscience; Virology.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Ariotti S., Hogenbirk M.A., Dijkgraaf F.E., Visser L.L., Hoekstra M.E., Song J.Y., Jacobs H., Haanen J.B., Schumacher T.N. T cell memory. Skin-resident memory CD8(+) T cells trigger a state of tissue-wide pathogen alert. Science. 2014;346:101–105. - PubMed
-
- Bengsch B., Spangenberg H.C., Kersting N., Neumann-Haefelin C., Panther E., von Weizsacker F., Blum H.E., Pircher H., Thimme R. Analysis of CD127 and KLRG1 expression on hepatitis C virus-specific CD8+ T cells reveals the existence of different memory T-cell subsets in the peripheral blood and liver. J. Virol. 2007;81:945–953. - PMC - PubMed
-
- Bird L. Lymphocyte responses: Hunker down with HOBIT and BLIMP1. Nat. Rev. Immunol. 2016;16:338–339. - PubMed
-
- Blais V., Rivest S. Effects of TNF-alpha and IFN-gamma on nitric oxide-induced neurotoxicity in the mouse brain. J. Immunol. 2004;172:7043–7052. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
