

Fungal evolution: diversity, taxonomy and phylogeny of the Fungi
- PMID: 31659870
- PMCID: PMC6899921
- DOI: 10.1111/brv.12550
Fungal evolution: diversity, taxonomy and phylogeny of the Fungi
Abstract
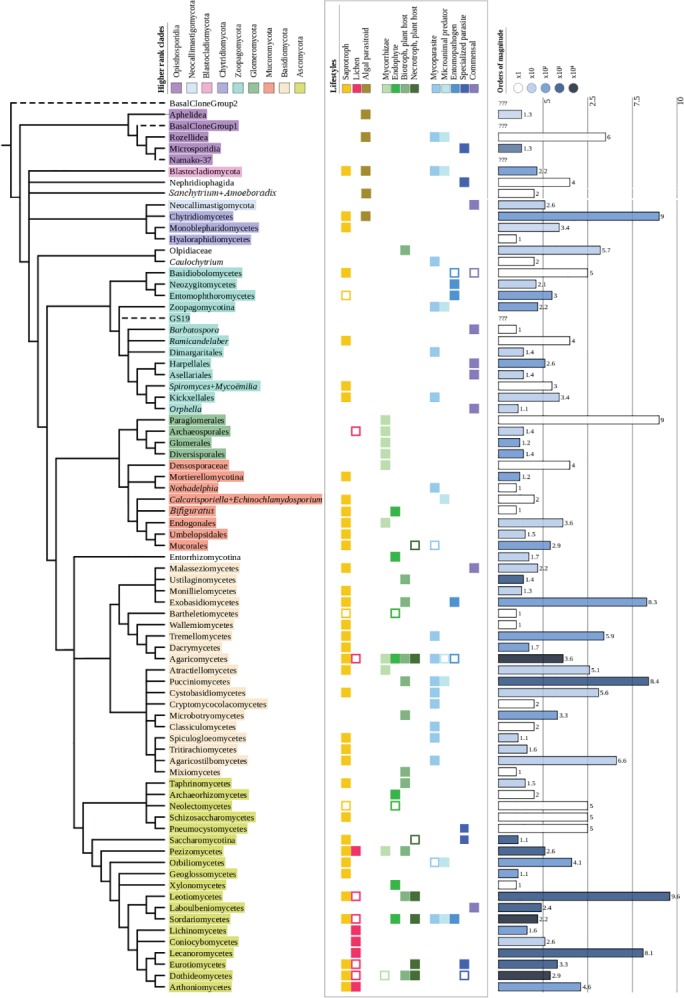
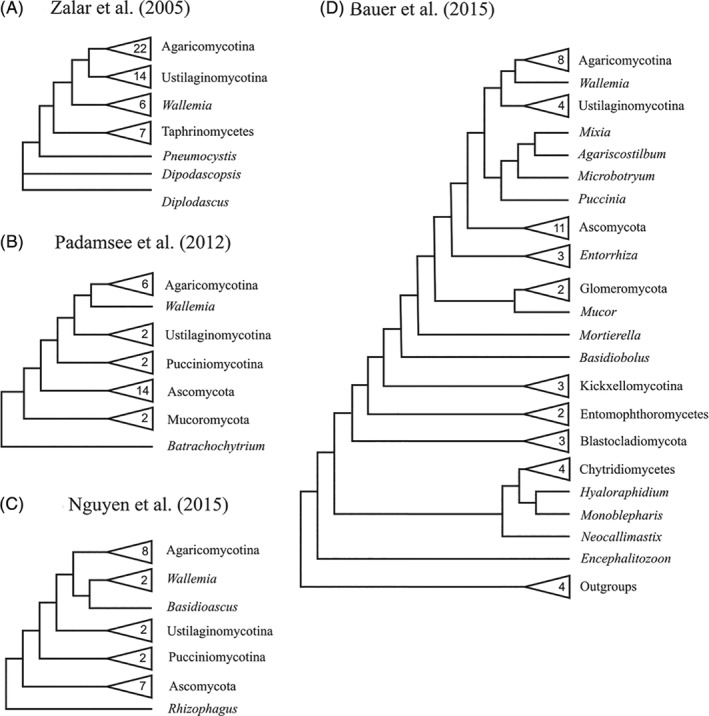
The fungal kingdom comprises a hyperdiverse clade of heterotrophic eukaryotes characterized by the presence of a chitinous cell wall, the loss of phagotrophic capabilities and cell organizations that range from completely unicellular monopolar organisms to highly complex syncitial filaments that may form macroscopic structures. Fungi emerged as a 'Third Kingdom', embracing organisms that were outside the classical dichotomy of animals versus vegetals. The taxonomy of this group has a turbulent history that is only now starting to be settled with the advent of genomics and phylogenomics. We here review the current status of the phylogeny and taxonomy of fungi, providing an overview of the main defined groups. Based on current knowledge, nine phylum-level clades can be defined: Opisthosporidia, Chytridiomycota, Neocallimastigomycota, Blastocladiomycota, Zoopagomycota, Mucoromycota, Glomeromycota, Basidiomycota and Ascomycota. For each group, we discuss their main traits and their diversity, focusing on the evolutionary relationships among the main fungal clades. We also explore the diversity and phylogeny of several groups of uncertain affinities and the main phylogenetic and taxonomical controversies and hypotheses in the field.
Keywords: Fungi; diversity; phylogenomics; phylogeny; taxonomy.
© 2019 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures



References
-
- Adl, S. M. , Simpson, A. G. , Lane, C. E. , Lukeš, J. , Bass, D. , Bowser, S. S. , Brown, M. , Burki, F. , Dunthorn, M. , Hampl, V. , Heiss, A. , Hoppenrath, M. , Lara, E. , Le Gall, L. , Lynn, D. H. , et al. (2012). The revised classification of eukaryotes. Journal of Eukaryotic Microbiology 59, 1–45. - PMC - PubMed
-
- Adl, S. M. , Bass, D. , Lane, C. E. , Lukeš, J. , Schoch, C. L. , Smirnov, A. , Agatha, S. , Berney, C. , Brown, M. W. , Burki, F. , Cárdenas, P. , Čepička, I. , Chistyakova, L. , del Campo, J. , Dunthorn, M. , et al. (2018). Revisions to the classification, nomenclature, and diversity of eukaryotes. Journal of Eukaryotic Microbiology 66, 4–119. - PMC - PubMed
-
- Ahrendt, S. R. , Quandt, C. A. , Ciobanu, D. , Clum, A. , Salamov, A. , Andreopoulos, B. , Cheng, J.‐F. , Woyke, T. , Pelin, A. , Henrissat, B. , Reynolds, N. K. , Benny, G. L. , Smith, M. E. , James, T. Y. , Grigoriev, I. V. , et al. (2018). Leveraging single‐cell genomics to expand the fungal tree of life. Nature Microbiology 3, 1417–1428. - PMC - PubMed
-
- Aime, M. C. , Matheny, P. B. , Henk, D. A. , Frieders, E. M. , Nilsson, R. H. , Piepenbring, M. , Mclaughlin, D. J. , Szabo, L. J. , Begerow, D. , Sampaio, J. P. , Bauer, R. , Weiß, M. , Oberwinkler, F. & Hibbett, D. (2006). An overview of the higher level classification of Pucciniomycotina based on combined analyses of nuclear large and small subunit rDNA sequences. Mycologia 98, 896–905. - PubMed
-
- Aime, M. C. , Toome, M. & McLaughlin, D. J. (2014). 10 Pucciniomycotina In Systematics and Evolution (eds Karl Esser), pp. 271–294. Springer, Berlin Heidelberg.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
