

Molecular Links between Central Obesity and Breast Cancer
- PMID: 31661891
- PMCID: PMC6862548
- DOI: 10.3390/ijms20215364
Molecular Links between Central Obesity and Breast Cancer
Abstract
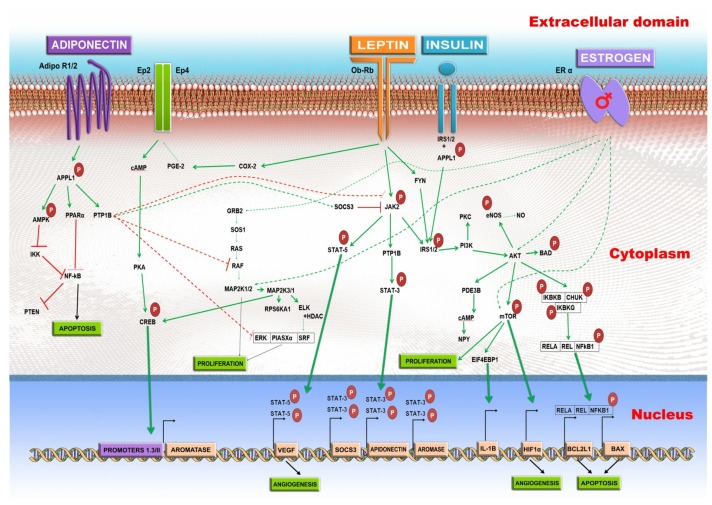
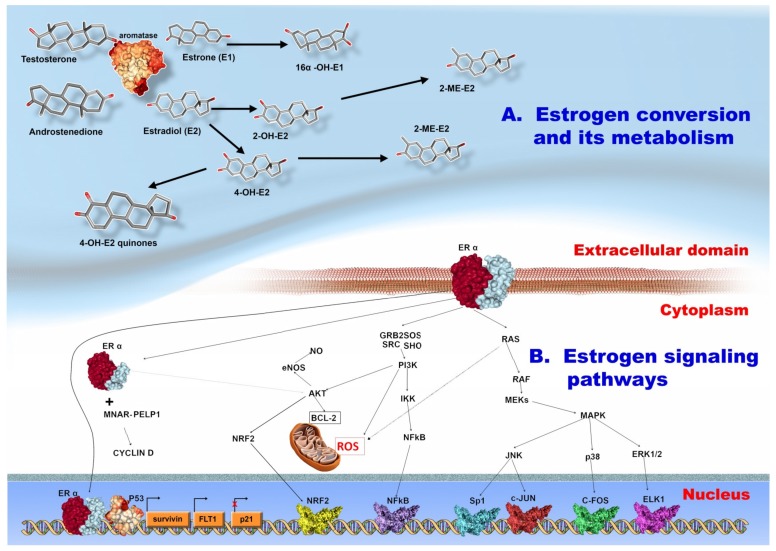
: Worldwide, breast cancer (BC) is the most common malignancy in women, in regard to incidence and mortality. In recent years, the negative role of obesity during BC development and progression has been made abundantly clear in several studies. However, the distribution of body fat may be more important to analyze than the overall body weight. In our review of literature, we reported some key findings regarding the role of obesity in BC development, but focused more on central adiposity. Firstly, the adipose microenvironment in obese people bears many similarities with the tumor microenvironment, in respect to associated cellular composition, chronic low-grade inflammation, and high ratio of reactive oxygen species to antioxidants. Secondly, the adipose tissue functions as an endocrine organ, which in obese people produces a high level of tumor-promoting hormones, such as leptin and estrogen, and a low level of the tumor suppressor hormone, adiponectin. As follows, in BC this leads to the activation of oncogenic signaling pathways: NFκB, JAK, STAT3, AKT. Moreover, overall obesity, but especially central obesity, promotes a systemic and local low grade chronic inflammation that further stimulates the increase of tumor-promoting oxidative stress. Lastly, there is a constant exchange of information between BC cells and adipocytes, mediated especially by extracellular vesicles, and which changes the transcription profile of both cell types to an oncogenic one with the help of regulatory non-coding RNAs.
Keywords: abdominal fat; adiponectin; breast cancer; exosomes; hormone dependency; leptin; menopause; miRNA; obesity.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
