

Cryo-electron microscopy structure and analysis of the P-Rex1-Gβγ signaling scaffold
- PMID: 31663027
- PMCID: PMC6795519
- DOI: 10.1126/sciadv.aax8855
Cryo-electron microscopy structure and analysis of the P-Rex1-Gβγ signaling scaffold
Abstract
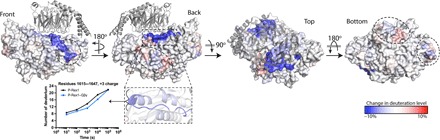
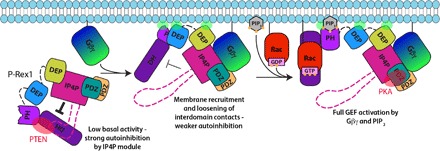
PIP3-dependent Rac exchanger 1 (P-Rex1) is activated downstream of G protein-coupled receptors to promote neutrophil migration and metastasis. The structure of more than half of the enzyme and its regulatory G protein binding site are unknown. Our 3.2 Å cryo-EM structure of the P-Rex1-Gβγ complex reveals that the carboxyl-terminal half of P-Rex1 adopts a complex fold most similar to those of Legionella phosphoinositide phosphatases. Although catalytically inert, the domain coalesces with a DEP domain and two PDZ domains to form an extensive docking site for Gβγ. Hydrogen-deuterium exchange mass spectrometry suggests that Gβγ binding induces allosteric changes in P-Rex1, but functional assays indicate that membrane localization is also required for full activation. Thus, a multidomain assembly is key to the regulation of P-Rex1 by Gβγ and the formation of a membrane-localized scaffold optimized for recruitment of other signaling proteins such as PKA and PTEN.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
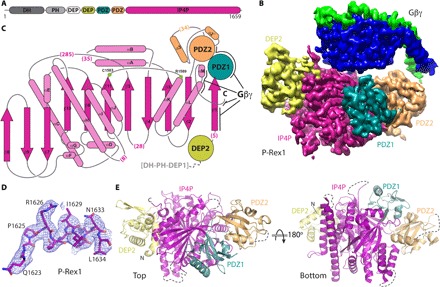
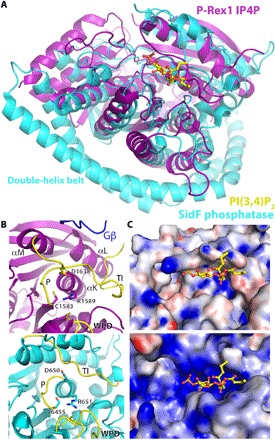
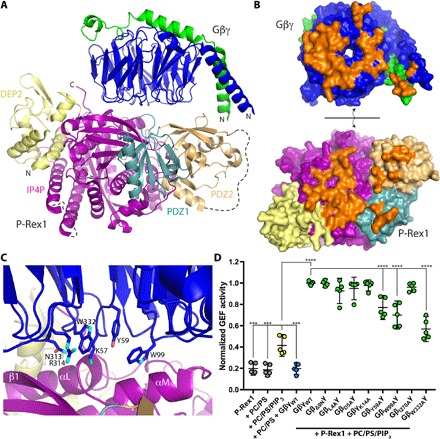
References
-
- Montero J. C., Seoane S., Ocaña A., Pandiella A., P-Rex1 participates in Neuregulin-ErbB signal transduction and its expression correlates with patient outcome in breast cancer. Oncogene 30, 1059–1071 (2011). - PubMed
-
- Lindsay C. R., Lawn S., Campbell A. D., Faller W. J., Rambow F., Mort R. L., Timpson P., Li A., Cammareri P., Ridgway R. A., Morton J. P., Doyle B., Hegarty S., Rafferty M., Murphy I. G., McDermott E. W., Sheahan K., Pedone K., Finn A. J., Groben P. A., Thomas N. E., Hao H., Carson C., Norman J. C., Machesky L. M., Gallagher W. M., Jackson I. J., van Kempen L., Beermann F., der C., Larue L., Welch H. C., Ozanne B. W., Sansom O. J., P-Rex1 is required for efficient melanoblast migration and melanoma metastasis. Nat. Commun. 2, 555–559 (2011). - PMC - PubMed
-
- Sosa M. S., Lopez-Haber C., Yang C., Wang H. B., Lemmon M. A., Busillo J. M., Luo J., Benovic J. L., Klein-Szanto A., Yagi H., Gutkind J. S., Parsons R. E., Kazanietz M. G., Identification of the Rac-GEF P-Rex1 as an essential mediator of ErbB signaling in breast cancer. Mol. Cell 40, 877–892 (2010). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
