

Large-scale genomic sequence data resolve the deepest divergences in the legume phylogeny and support a near-simultaneous evolutionary origin of all six subfamilies
- PMID: 31665814
- PMCID: PMC6972672
- DOI: 10.1111/nph.16290
Large-scale genomic sequence data resolve the deepest divergences in the legume phylogeny and support a near-simultaneous evolutionary origin of all six subfamilies
Abstract
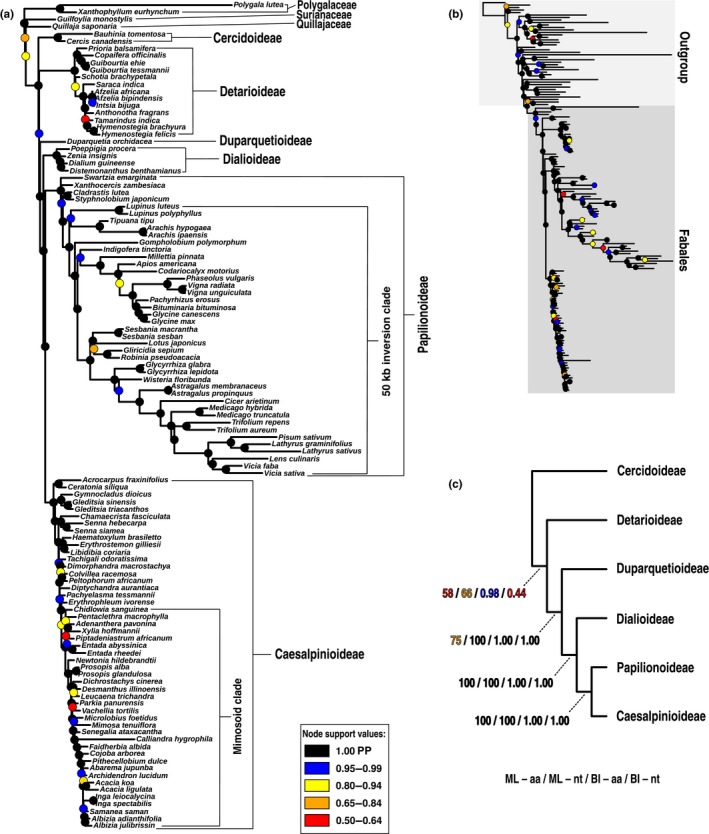
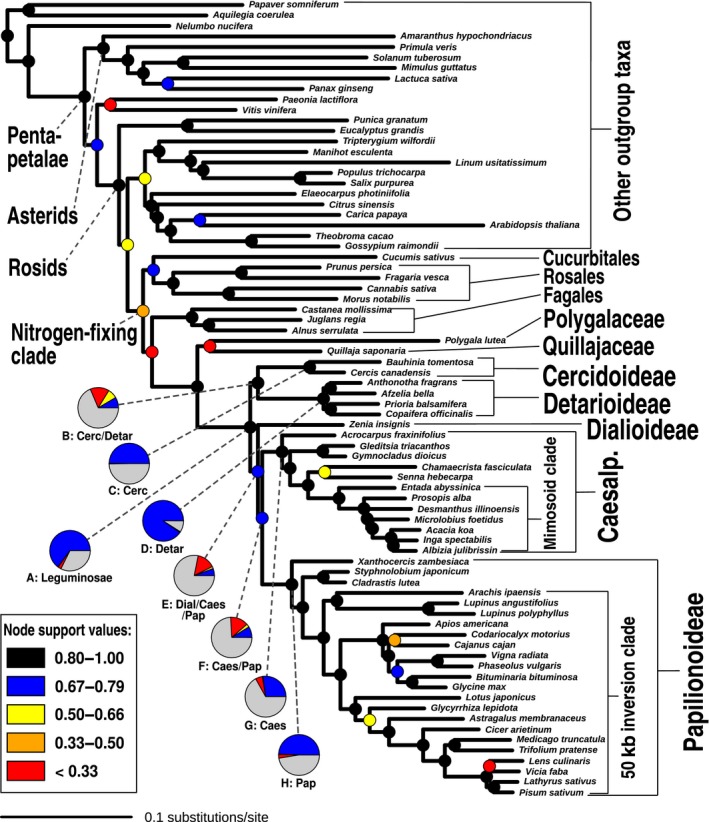
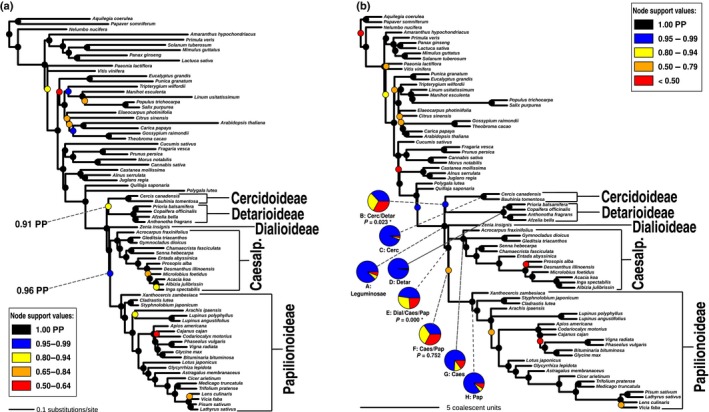
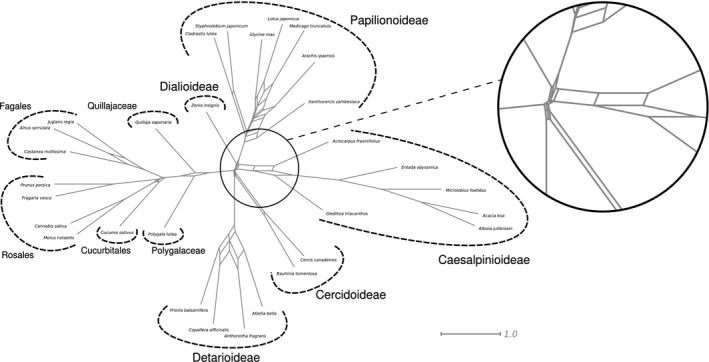
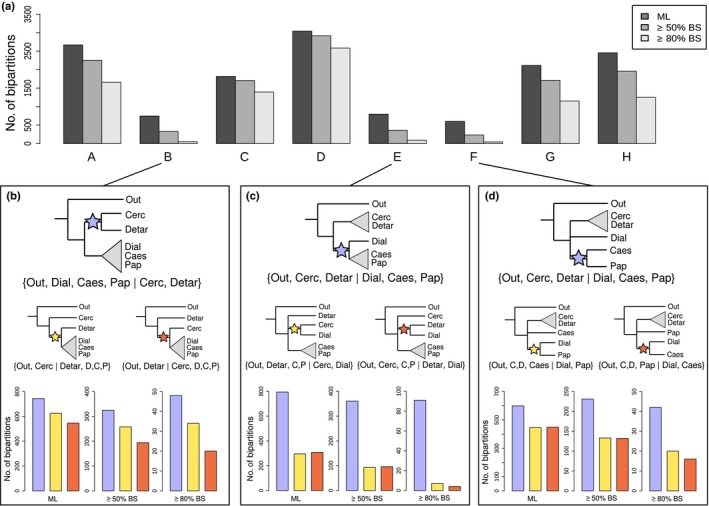
Phylogenomics is increasingly used to infer deep-branching relationships while revealing the complexity of evolutionary processes such as incomplete lineage sorting, hybridization/introgression and polyploidization. We investigate the deep-branching relationships among subfamilies of the Leguminosae (or Fabaceae), the third largest angiosperm family. Despite their ecological and economic importance, a robust phylogenetic framework for legumes based on genome-scale sequence data is lacking. We generated alignments of 72 chloroplast genes and 7621 homologous nuclear-encoded proteins, for 157 and 76 taxa, respectively. We analysed these with maximum likelihood, Bayesian inference, and a multispecies coalescent summary method, and evaluated support for alternative topologies across gene trees. We resolve the deepest divergences in the legume phylogeny despite lack of phylogenetic signal across all chloroplast genes and the majority of nuclear genes. Strongly supported conflict in the remainder of nuclear genes is suggestive of incomplete lineage sorting. All six subfamilies originated nearly simultaneously, suggesting that the prevailing view of some subfamilies as 'basal' or 'early-diverging' with respect to others should be abandoned, which has important implications for understanding the evolution of legume diversity and traits. Our study highlights the limits of phylogenetic resolution in relation to rapid successive speciation.
Keywords: Fabaceae; Leguminosae; gene tree conflict; incomplete lineage sorting; lack of phylogenetic signal; phylogenomics.
© 2019 The Authors. New Phytologist © 2019 New Phytologist Trust.
Figures

References
-
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, Pyshkin AV, Sirotkin AV, Vyahhi N, Tesler G, Alekseyev MA, Pevzner PA. 2012. SPAdes: a new genome assembly algorithm and its applications to single‐cell sequencing. Journal of Computational Biology 19: 455–477. - PMC - PubMed
-
- Bruneau A, Mercure M, Lewis GP, Herendeen PS. 2008. Phylogenetic patterns and diversification in the caesalpinioid legumes. Botany 86: 697–718.
-
- Bruneau A, Klitgaard BB, Prenner G, Fougere‐Danezan M, Tucker SC. 2014. Floral evolution in the Detarieae (Leguminosae): phylogenetic evidence for labile floral development in an early‐diverging legume lineage. International Journal of Plant Sciences 175: 392–417.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
