

Mechanisms of soft tissue and protein preservation in Tyrannosaurus rex
- PMID: 31666554
- PMCID: PMC6821828
- DOI: 10.1038/s41598-019-51680-1
Mechanisms of soft tissue and protein preservation in Tyrannosaurus rex
Abstract
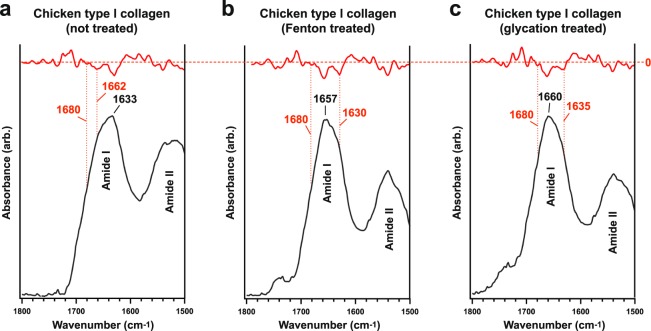
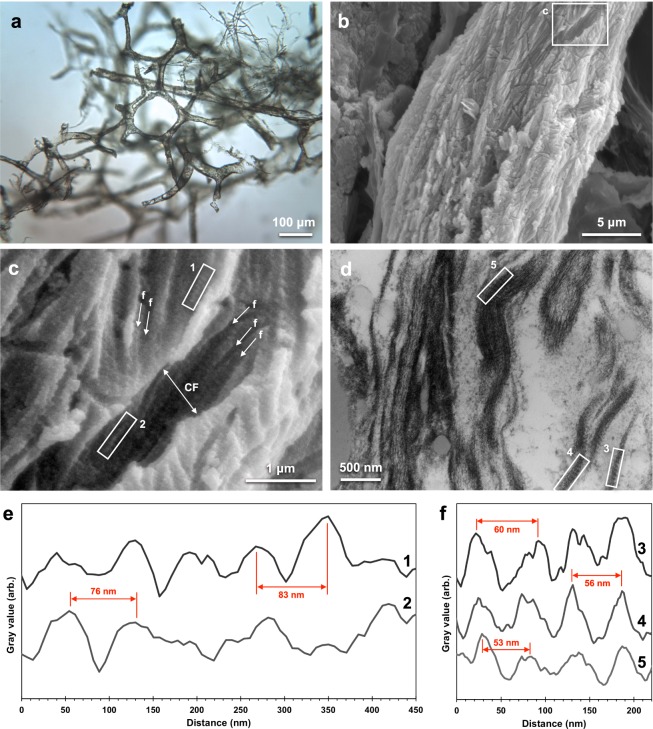
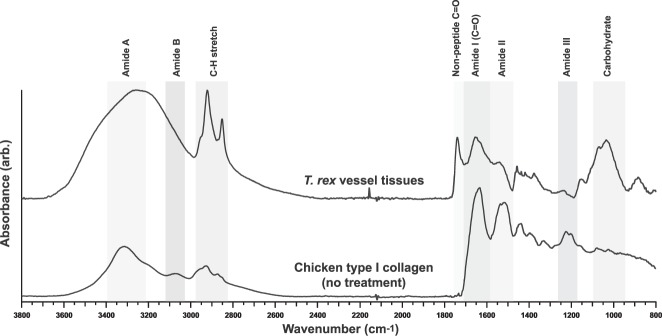
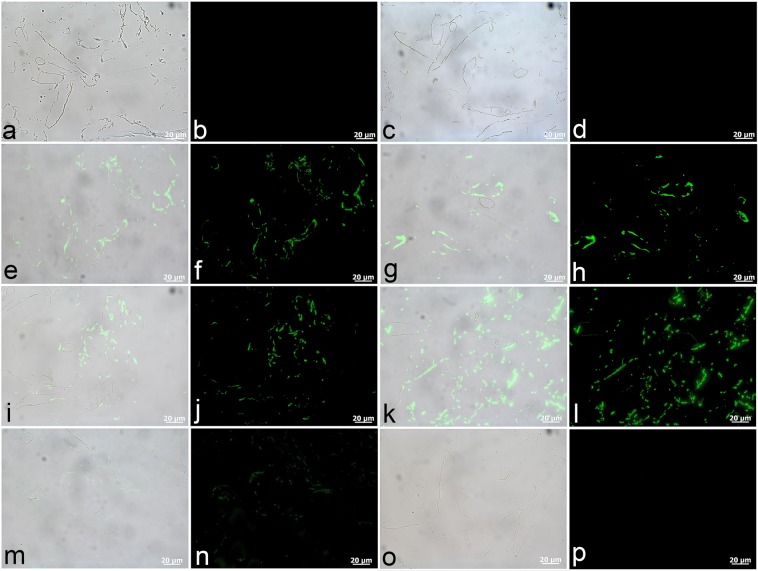
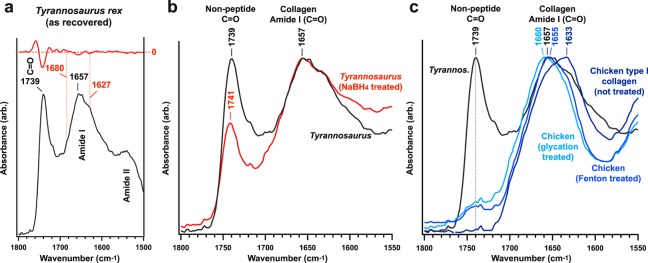
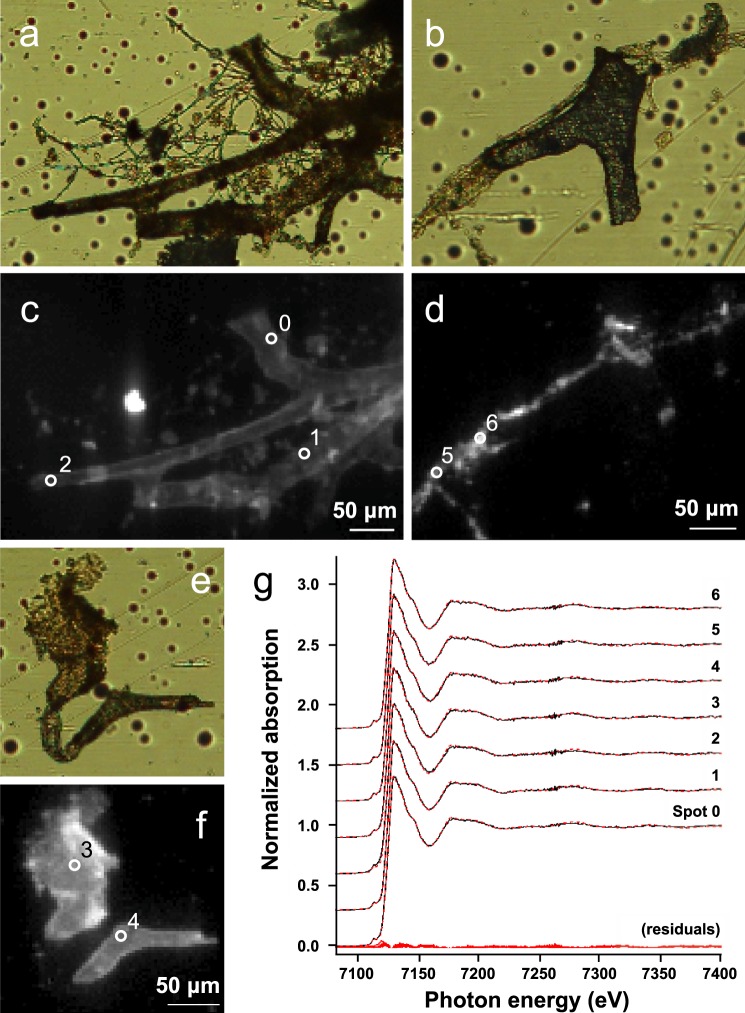
The idea that original soft tissue structures and the native structural proteins comprising them can persist across geological time is controversial, in part because rigorous and testable mechanisms that can occur under natural conditions, resulting in such preservation, have not been well defined. Here, we evaluate two non-enzymatic structural protein crosslinking mechanisms, Fenton chemistry and glycation, for their possible contribution to the preservation of blood vessel structures recovered from the cortical bone of a Tyrannosaurus rex (USNM 555000 [formerly, MOR 555]). We demonstrate the endogeneity of the fossil vessel tissues, as well as the presence of type I collagen in the outermost vessel layers, using imaging, diffraction, spectroscopy, and immunohistochemistry. Then, we use data derived from synchrotron FTIR studies of the T. rex vessels to analyse their crosslink character, with comparison against two non-enzymatic Fenton chemistry- and glycation-treated extant chicken samples. We also provide supporting X-ray microprobe analyses of the chemical state of these fossil tissues to support our conclusion that non-enzymatic crosslinking pathways likely contributed to stabilizing, and thus preserving, these T. rex vessels. Finally, we propose that these stabilizing crosslinks could play a crucial role in the preservation of other microvascular tissues in skeletal elements from the Mesozoic.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
