

Rewiring carbon metabolism in yeast for high level production of aromatic chemicals
- PMID: 31672987
- PMCID: PMC6823513
- DOI: 10.1038/s41467-019-12961-5
Rewiring carbon metabolism in yeast for high level production of aromatic chemicals
Abstract
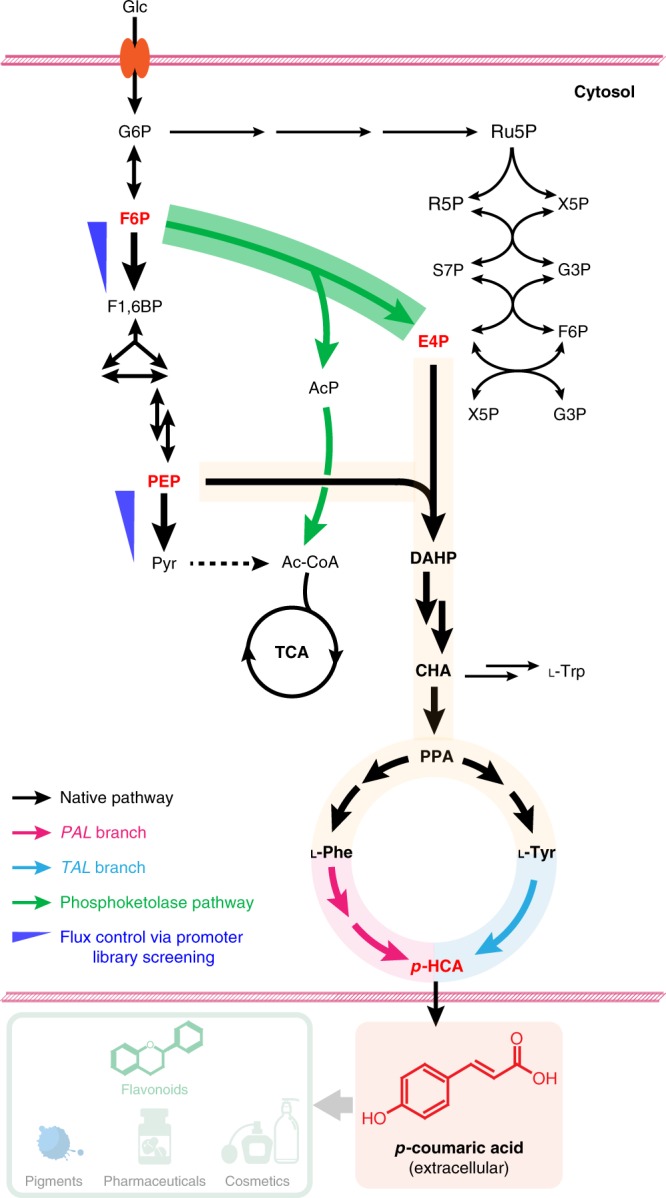
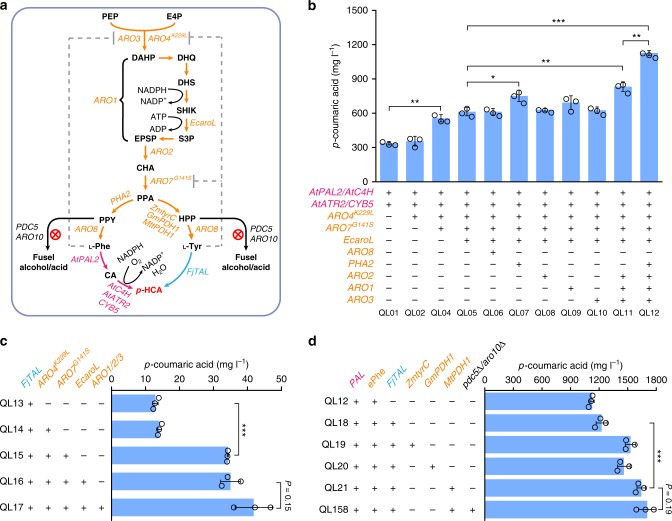
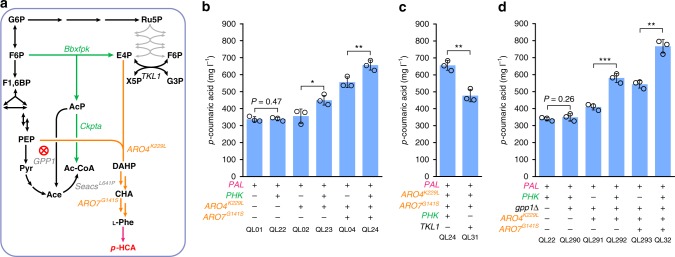
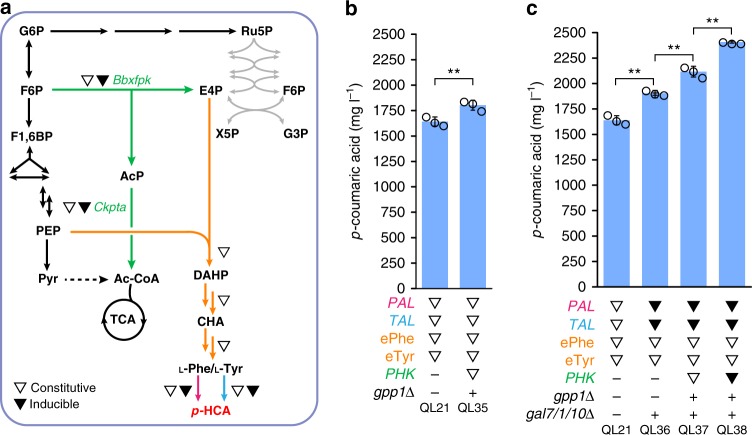
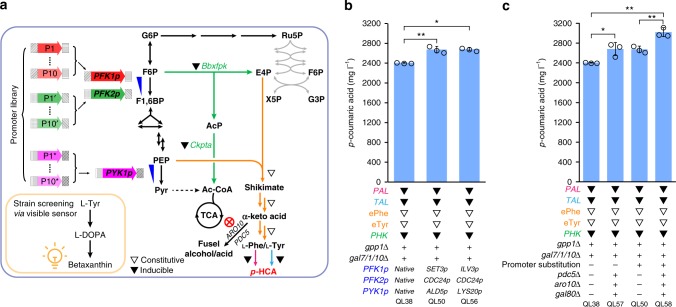
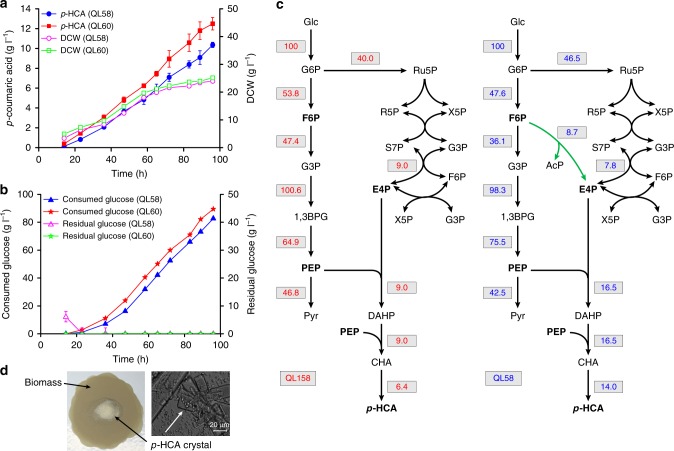
The production of bioactive plant compounds using microbial hosts is considered a safe, cost-competitive and scalable approach to their production. However, microbial production of some compounds like aromatic amino acid (AAA)-derived chemicals, remains an outstanding metabolic engineering challenge. Here we present the construction of a Saccharomyces cerevisiae platform strain able to produce high levels of p-coumaric acid, an AAA-derived precursor for many commercially valuable chemicals. This is achieved through engineering the AAA biosynthesis pathway, introducing a phosphoketalose-based pathway to divert glycolytic flux towards erythrose 4-phosphate formation, and optimizing carbon distribution between glycolysis and the AAA biosynthesis pathway by replacing the promoters of several important genes at key nodes between these two pathways. This results in a maximum p-coumaric acid titer of 12.5 g L-1 and a maximum yield on glucose of 154.9 mg g-1.
Conflict of interest statement
Q.L.L., J.N., and Y.C. are inventors on a pending patent application on utilization of phosphoketolase for the production of aromatic amino acid-derived products (EP18207629.9). All other authors declare no competing interests.
Figures
Similar articles
-
Rewiring Central Carbon Metabolism Ensures Increased Provision of Acetyl-CoA and NADPH Required for 3-OH-Propionic Acid Production.ACS Synth Biol. 2020 Dec 18;9(12):3236-3244. doi: 10.1021/acssynbio.0c00264. Epub 2020 Nov 13. ACS Synth Biol. 2020. PMID: 33186034
-
Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis.Metab Eng. 2015 Sep;31:181-8. doi: 10.1016/j.ymben.2015.08.003. Epub 2015 Aug 18. Metab Eng. 2015. PMID: 26292030
-
Metabolic engineering and transcriptomic analysis of Saccharomyces cerevisiae producing p-coumaric acid from xylose.Microb Cell Fact. 2019 Nov 5;18(1):191. doi: 10.1186/s12934-019-1244-4. Microb Cell Fact. 2019. PMID: 31690329 Free PMC article.
-
Pathway engineering for the production of heterologous aromatic chemicals and their derivatives in Saccharomyces cerevisiae: bioconversion from glucose.FEMS Yeast Res. 2017 Jun 1;17(4). doi: 10.1093/femsyr/fox035. FEMS Yeast Res. 2017. PMID: 28582489 Review.
-
Yeast factories for the production of aromatic compounds: from building blocks to plant secondary metabolites.J Ind Microbiol Biotechnol. 2016 Nov;43(11):1611-1624. doi: 10.1007/s10295-016-1824-9. Epub 2016 Aug 31. J Ind Microbiol Biotechnol. 2016. PMID: 27581441 Review.
Cited by
-
Dietary Supplementation With Chlorogenic Acid Derived From Lonicera macranthoides Hand-Mazz Improves Meat Quality and Muscle Fiber Characteristics of Finishing Pigs via Enhancement of Antioxidant Capacity.Front Physiol. 2021 Apr 20;12:650084. doi: 10.3389/fphys.2021.650084. eCollection 2021. Front Physiol. 2021. PMID: 33959038 Free PMC article.
-
Combining metabolite doping and metabolic engineering to improve 2-phenylethanol production by engineered cyanobacteria.Front Bioeng Biotechnol. 2022 Sep 20;10:1005960. doi: 10.3389/fbioe.2022.1005960. eCollection 2022. Front Bioeng Biotechnol. 2022. PMID: 36204466 Free PMC article.
-
Metabolic Engineering of Saccharomyces cerevisiae for Rosmarinic Acid Production.ACS Synth Biol. 2020 Aug 21;9(8):1978-1988. doi: 10.1021/acssynbio.0c00048. Epub 2020 Jul 27. ACS Synth Biol. 2020. PMID: 32589831 Free PMC article.
-
A DNA assembly toolkit to unlock the CRISPR/Cas9 potential for metabolic engineering.Commun Biol. 2023 Aug 18;6(1):858. doi: 10.1038/s42003-023-05202-5. Commun Biol. 2023. PMID: 37596335 Free PMC article.
-
Saccharomyces cerevisiae as host for the recombinant production of polyketides and nonribosomal peptides.Microb Cell Fact. 2021 Aug 19;20(1):161. doi: 10.1186/s12934-021-01650-y. Microb Cell Fact. 2021. PMID: 34412657 Free PMC article. Review.
References
-
- Kallscheuer N, Classen T, Drepper T, Marienhagen J. Production of plant metabolites with applications in the food industry using engineered microorganisms. Curr. Opin. Biotechnol. 2019;56:7–17. - PubMed
-
- Ehrenworth AM, Peralta-Yahya P. Accelerating the semisynthesis of alkaloid-based drugs through metabolic engineering. Nat. Chem. Biol. 2017;13:249–258. - PubMed
-
- Newman DJ, Cragg GM. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016;79:629–661. - PubMed
-
- Chemler JA, Koffas MA. Metabolic engineering for plant natural product biosynthesis in microbes. Curr. Opin. Biotechnol. 2008;19:597–605. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
