CEH-60/PBX regulates vitellogenesis and cuticle permeability through intestinal interaction with UNC-62/MEIS in Caenorhabditis elegans
- PMID: 31675356
- PMCID: PMC6824563
- DOI: 10.1371/journal.pbio.3000499
CEH-60/PBX regulates vitellogenesis and cuticle permeability through intestinal interaction with UNC-62/MEIS in Caenorhabditis elegans
Abstract
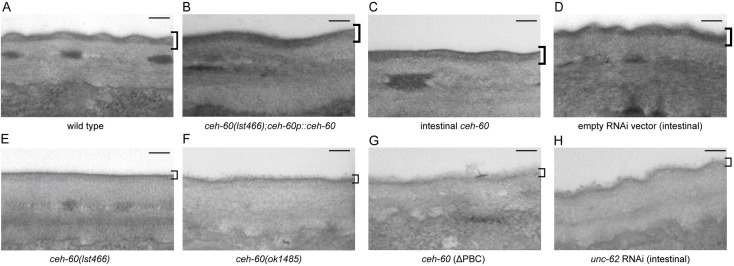
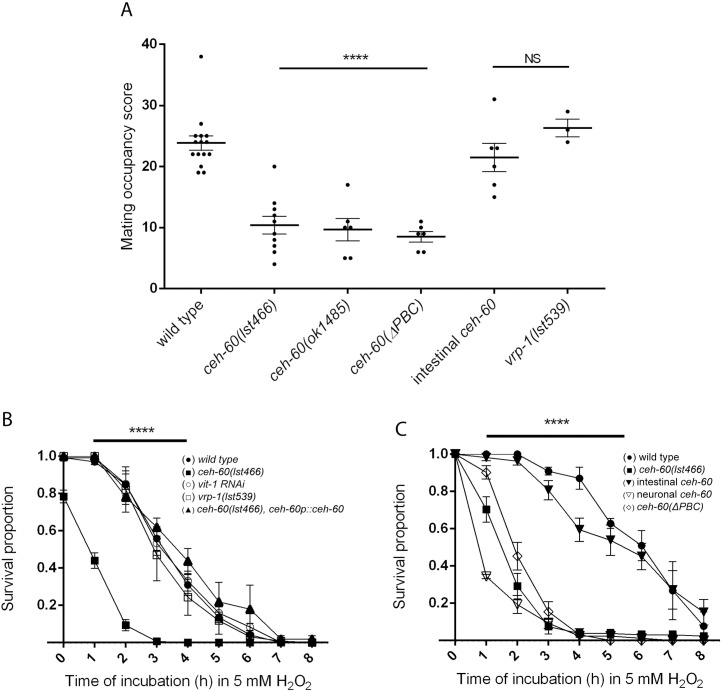
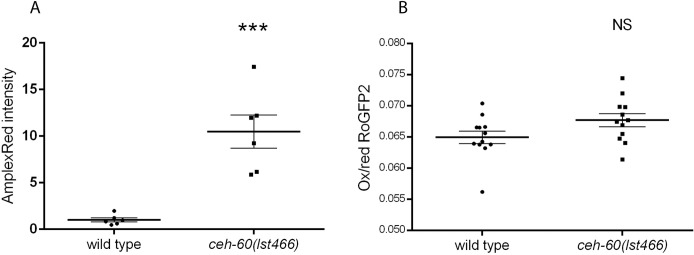
The onset of sexual maturity involves dramatic changes in physiology and gene expression in many animals. These include abundant yolk protein production in egg-laying species, an energetically costly process under extensive transcriptional control. Here, we used the model organism Caenorhabditis elegans to provide evidence for the spatiotemporally defined interaction of two evolutionarily conserved transcription factors, CEH-60/PBX and UNC-62/MEIS, acting as a gateway to yolk protein production. Via proteomics, bimolecular fluorescence complementation (BiFC), and biochemical and functional readouts, we show that this interaction occurs in the intestine of animals at the onset of sexual maturity and suffices to support the reproductive program. Our electron micrographs and functional assays provide evidence that intestinal PBX/MEIS cooperation drives another process that depends on lipid mobilization: the formation of an impermeable epicuticle. Without this lipid-rich protective layer, mutant animals are hypersensitive to exogenous oxidative stress and are poor partners for mating. Dedicated communication between the hypodermis and intestine in C. elegans likely supports these physiological outcomes, and we propose a fundamental role for the conserved PBX/MEIS interaction in multicellular signaling networks that rely on lipid homeostasis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures