

Alternatively spliced isoforms reveal a novel type of PTB domain in CCM2 protein
- PMID: 31676827
- PMCID: PMC6825194
- DOI: 10.1038/s41598-019-52386-0
Alternatively spliced isoforms reveal a novel type of PTB domain in CCM2 protein
Abstract
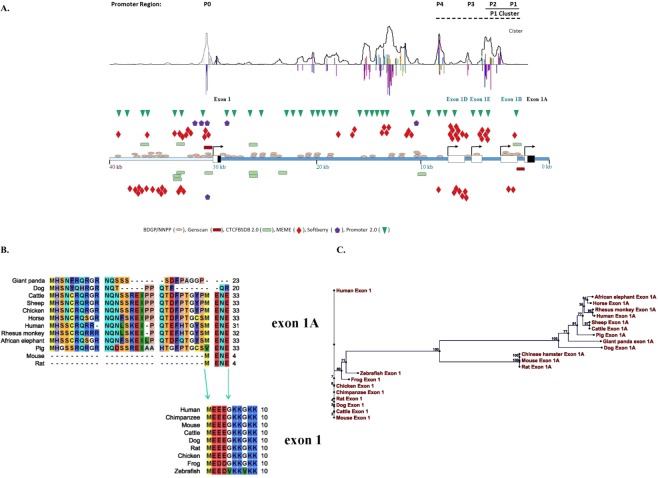
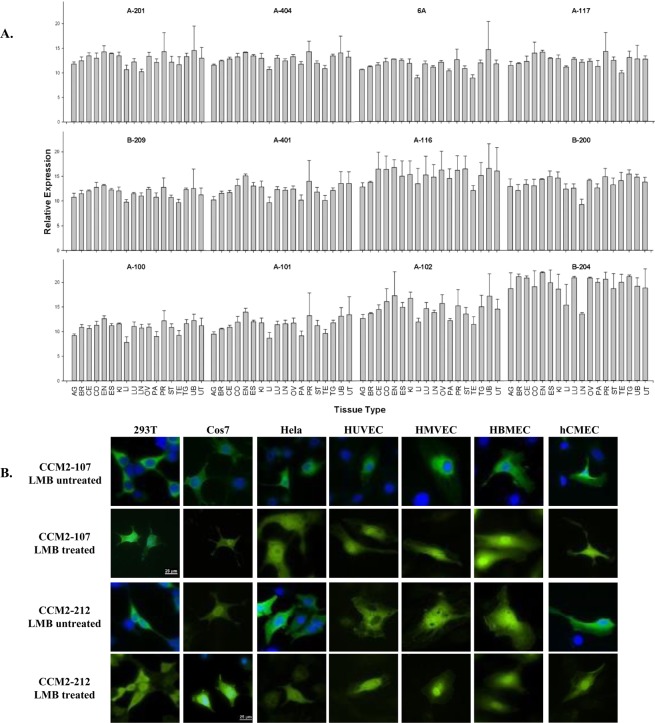
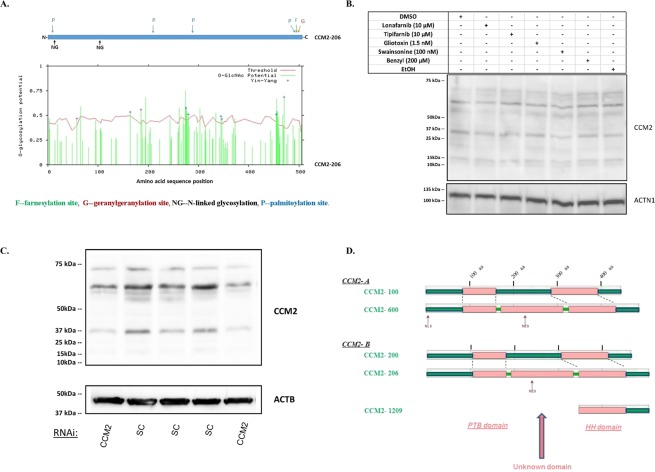
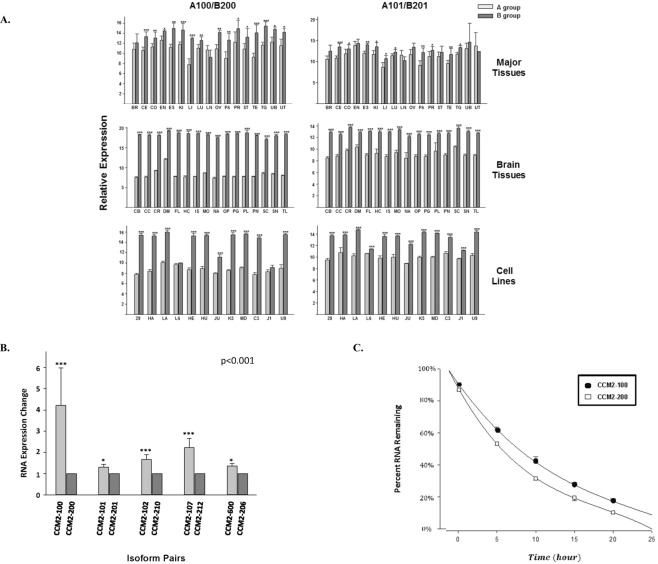
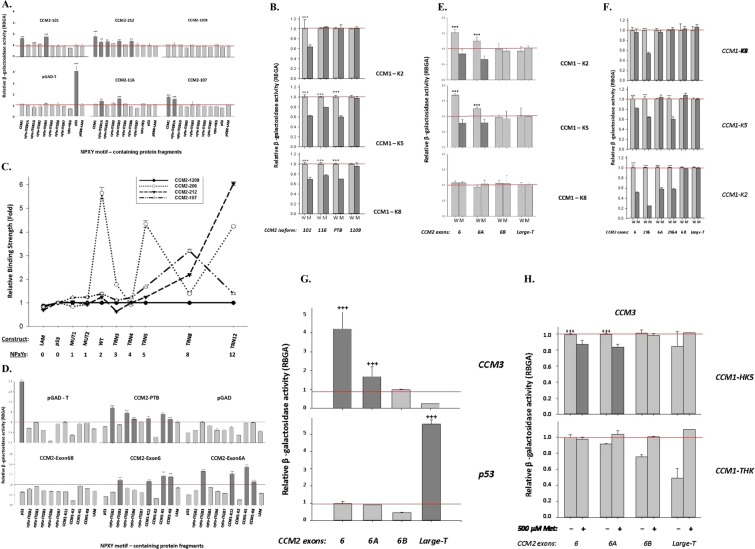
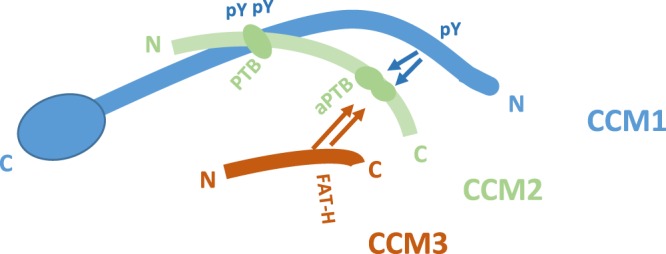
Cerebral cavernous malformations (CCMs) is a microvascular disorder in the central nervous system. Despite tremendous efforts, the causal genetic mutation in some CCM patients has not be identified, raising the possibility of an unknown CCM locus. The CCM2/MGC4607 gene has been identified as one of three known genes causing CCMs. In this report, we defined a total of 29 novel exons and 4 novel promoters in CCM2 genomic structure and subsequently identified a total of 50 new alternative spliced isoforms of CCM2 which eventually generated 22 novel protein isoforms. Genetic analysis of CCM2 isoforms revealed that the CCM2 isoforms can be classified into two groups based on their alternative promoters and alternative start codon exons. Our data demonstrated that CCM2 isoforms not only are specific in their subcellular compartmentation but also have distinct cellular expression patterns among various tissues and cells, indicating the pleiotropic cellular roles of CCM2 through their multiple isoforms. In fact, the complexity of the CCM2 genomic structure was reflected by the multiple layers of regulation of CCM2 expression patterns. At the transcriptional level, it is accomplished by alternative promoters, alternative splicing, and multiple transcriptional start sites and termination sites; while at the translational level, it is carried out with various cellular functions with a distinguishable CCM2 protein group pattern, specified abundance and composition of selective isoforms in a cell and tissue specific fashion. Through experimentation, we discovered a unique phosphotyrosine binding (PTB) domain, namely atypical phosphotyrosine binding (aPTB) domain. Some long CCM2 isoform proteins contain both classes of PTB domains, making them a dual PTB domain-containing protein. Both CCM1 and CCM3 can bind competitively to this aPTB domain, indicating CCM2 as the cornerstone for CCM signaling complex (CSC).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
