

The transfer of specific mitochondrial lipids and proteins to lipid droplets contributes to proteostasis upon stress and aging in the eukaryotic model system Saccharomyces cerevisiae
- PMID: 31676965
- PMCID: PMC7031196
- DOI: 10.1007/s11357-019-00103-0
The transfer of specific mitochondrial lipids and proteins to lipid droplets contributes to proteostasis upon stress and aging in the eukaryotic model system Saccharomyces cerevisiae
Abstract
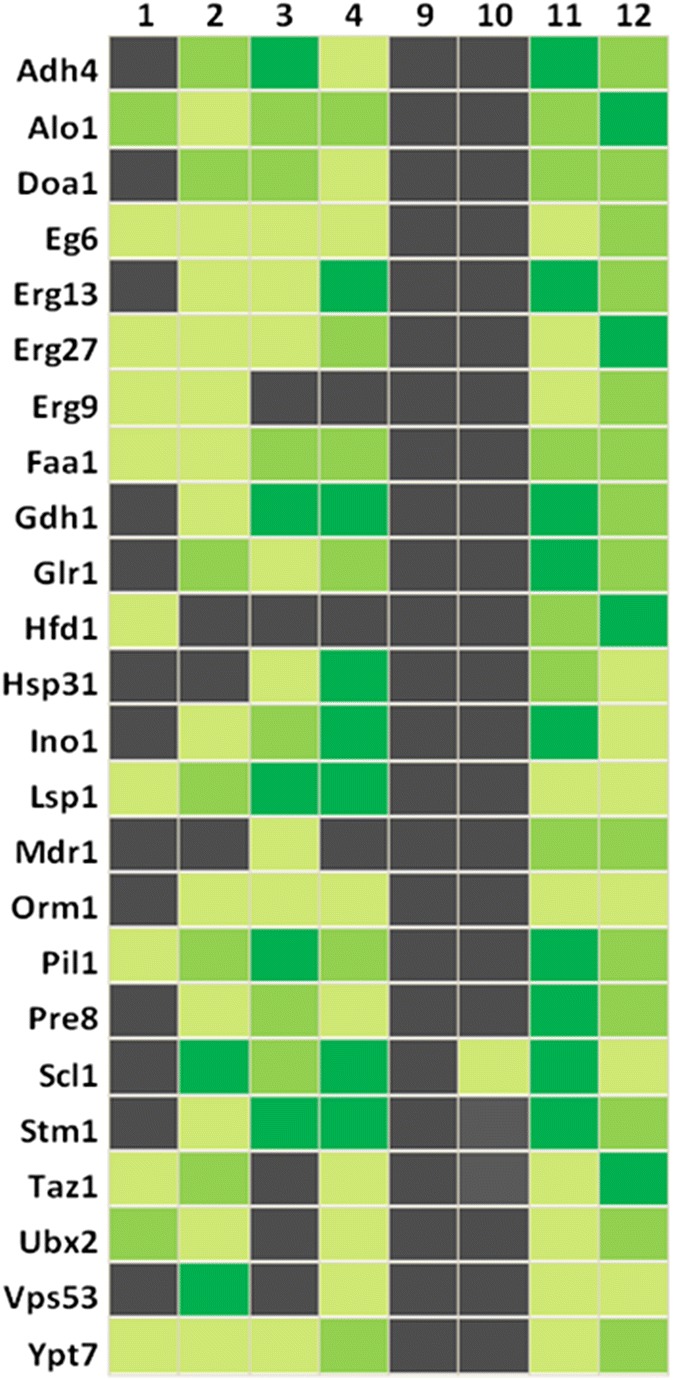
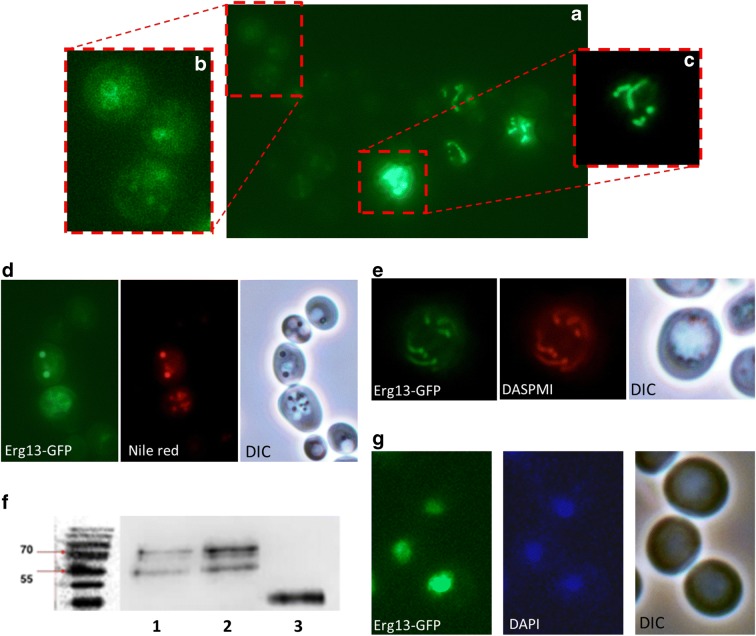
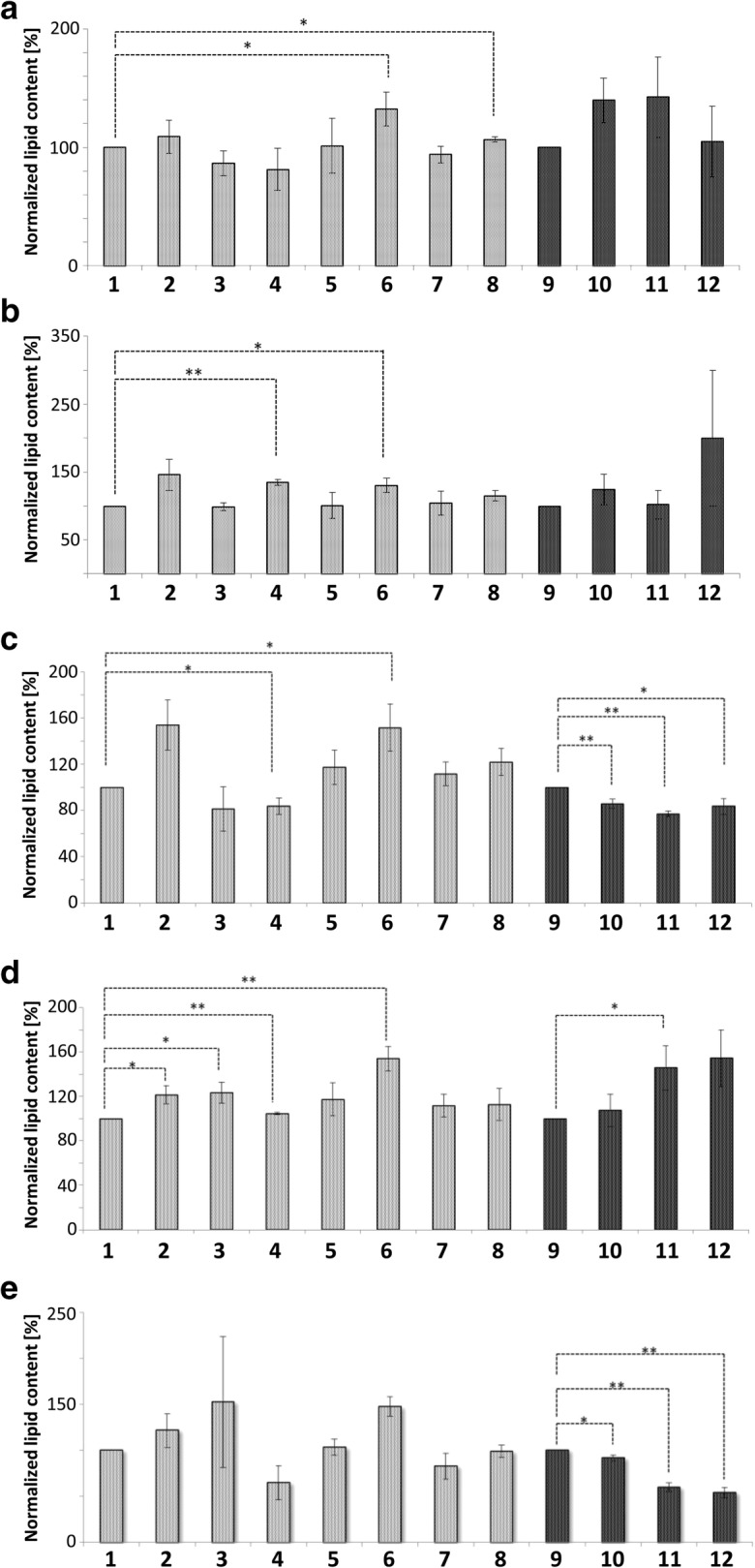
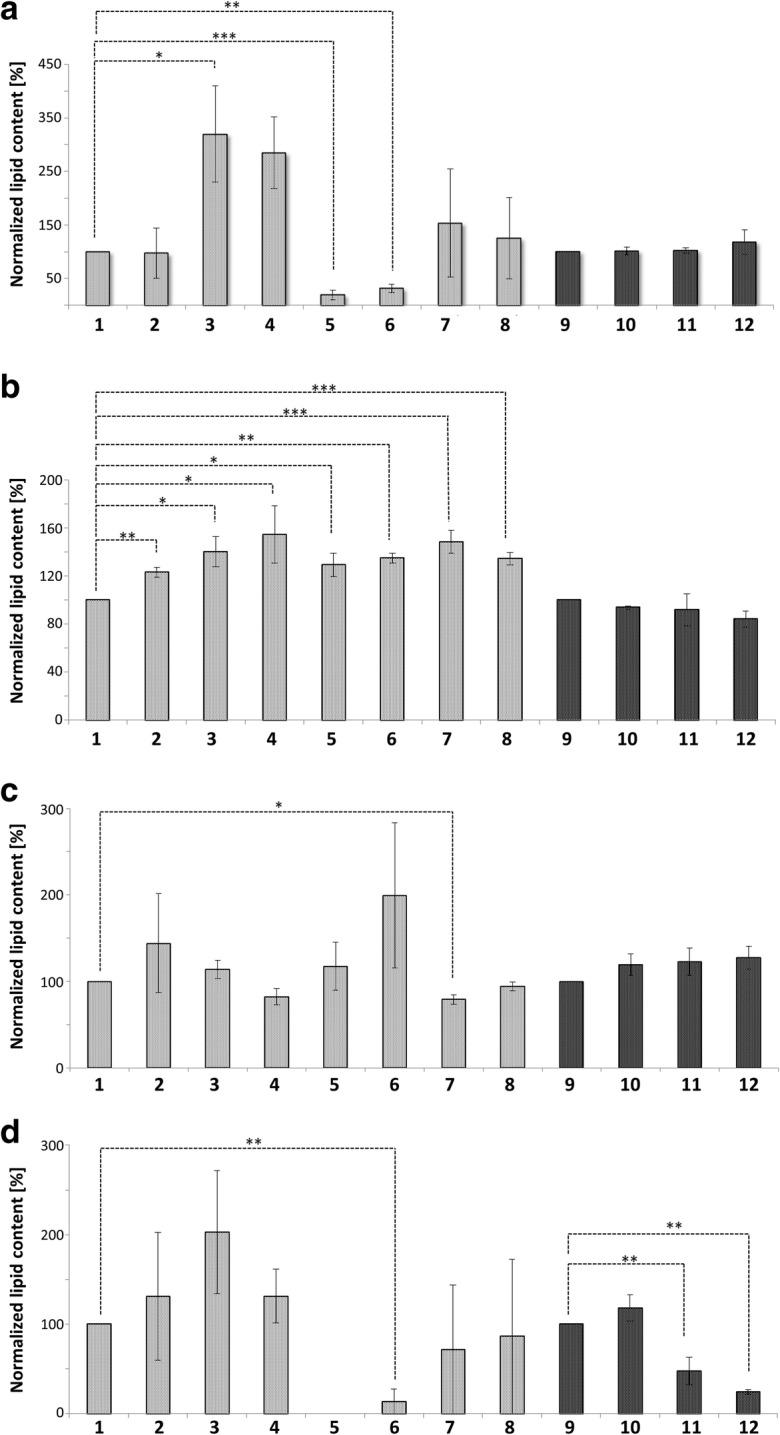
Originally Lipid droplets (LDs) were considered as being droplets for lipid storage only. Increasing evidence, however, demonstrates that LDs fulfill a pleiotropy of additional functions. Among them is the modulation of protein as well as lipid homeostasis. Under unfavorable pro-oxidative conditions, proteins can form aggregates which may exceed the overall proteolytic capacity of the proteasome. After stress termination LDs can adjust and support the removal of these aggregates. Additionally, LDs interact with mitochondria, specifically take over certain proteins and thus prevent apoptosis. LDs, which are loaded with these harmful proteins, are subsequently eliminated via lipophagy. Recently it was demonstrated that this autophagic process is a modulator of longevity. LDs do not only eliminate potentially dangerous proteins, but they are also able to prevent lipotoxicity by storing specific lipids. In the present study we used the model organism Saccharomyces cerevisiae to compare the proteome as well as lipidome of mitochondria and LDs under different conditions: replicative aging, stress and apoptosis. In this context we found an accumulation of proteins at LDs, supporting the role of LDs in proteostasis. Additionally, the composition of main lipid classes such as phosphatidylcholines, phosphatidylethanolamines, phosphatidylinositols, phosphatidylglycerols, triacylglycerols, ceramides, phosphatidic acids and ergosterol of LDs and mitochondria changed during stress conditions and aging.
Keywords: Lipid droplets; Lipidome; Mitochondria; Protein sink; Proteome; Replicative aging.
Figures
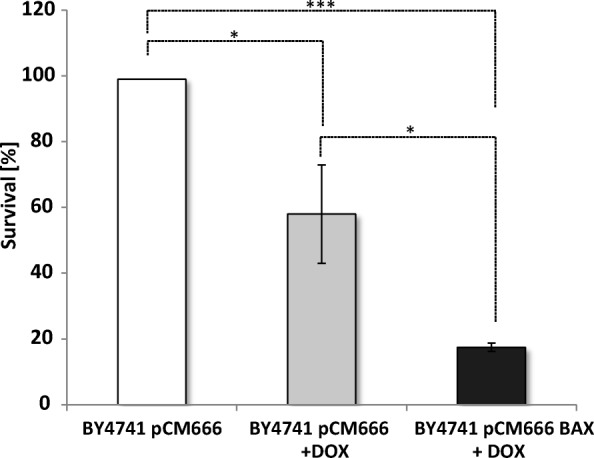
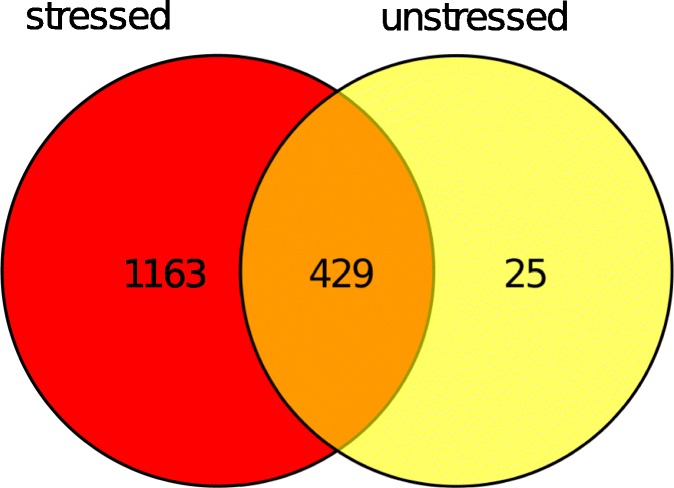
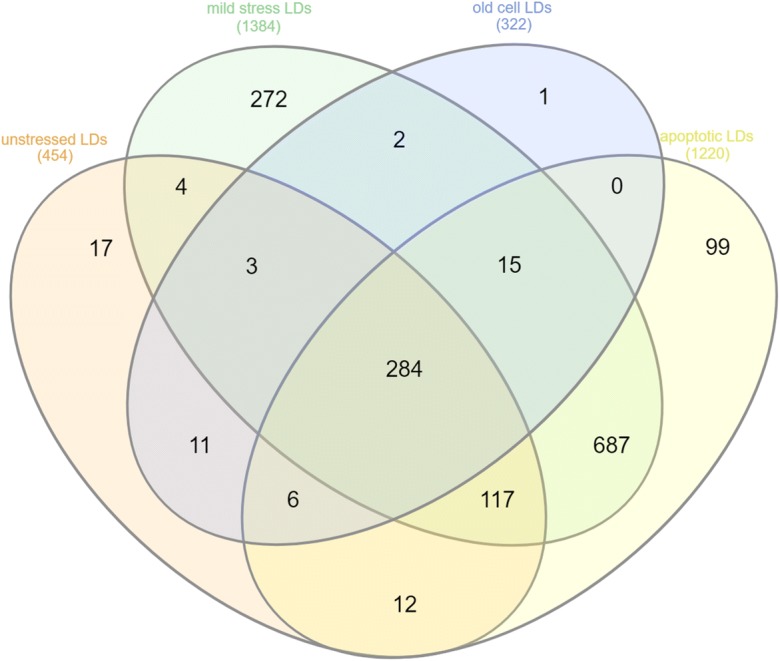
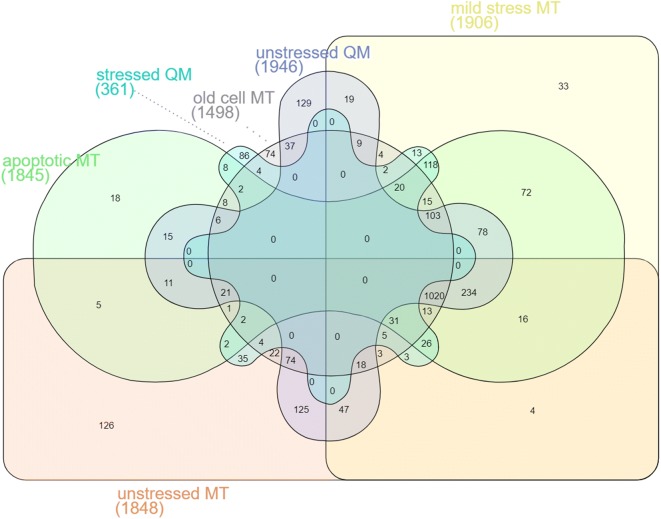
References
-
- Bischof J, Salzmann M, Streubel MK, Hasek J, Geltinger F, Duschl J, Bresgen N, Briza P, Haskova D, Lejskova R, Sopjani M, Richter K, Rinnerthaler M. Clearing the outer mitochondrial membrane from harmful proteins via lipid droplets. Cell Death Discov. 2017;3:17016. doi: 10.1038/cddiscovery.2017.16. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
