

An Ancient CFTR Ortholog Informs Molecular Evolution in ABC Transporters
- PMID: 31679858
- PMCID: PMC7665244
- DOI: 10.1016/j.devcel.2019.09.017
An Ancient CFTR Ortholog Informs Molecular Evolution in ABC Transporters
Abstract
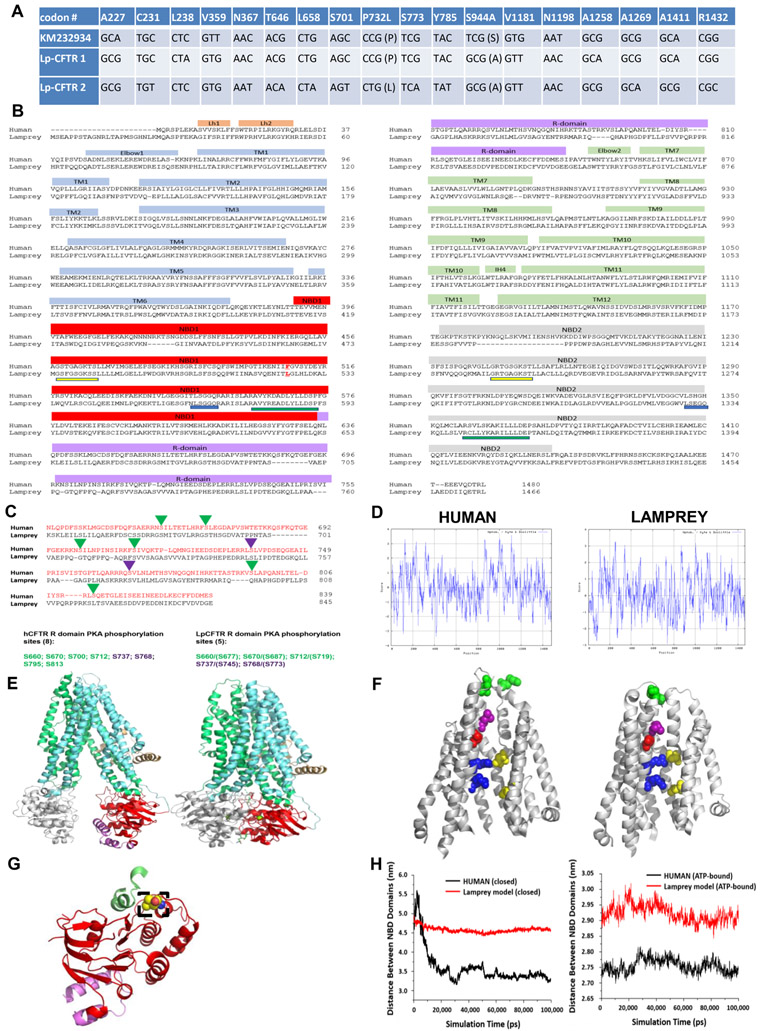
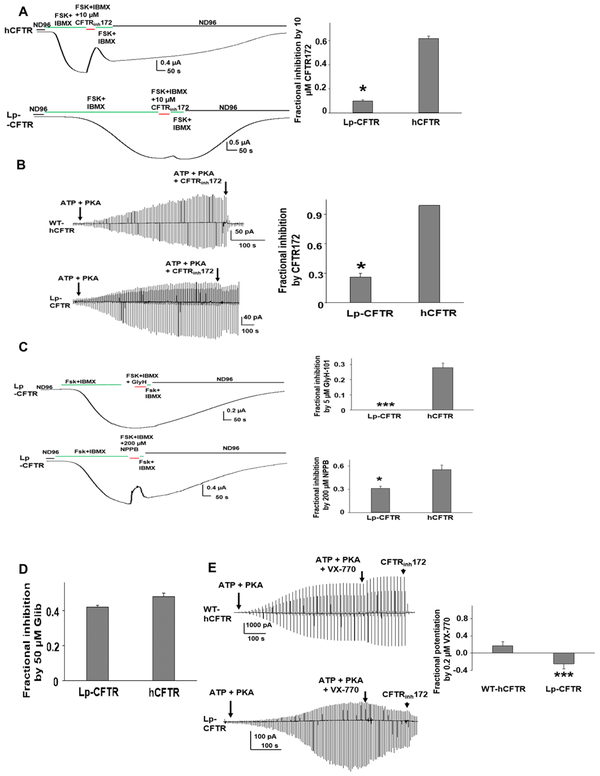
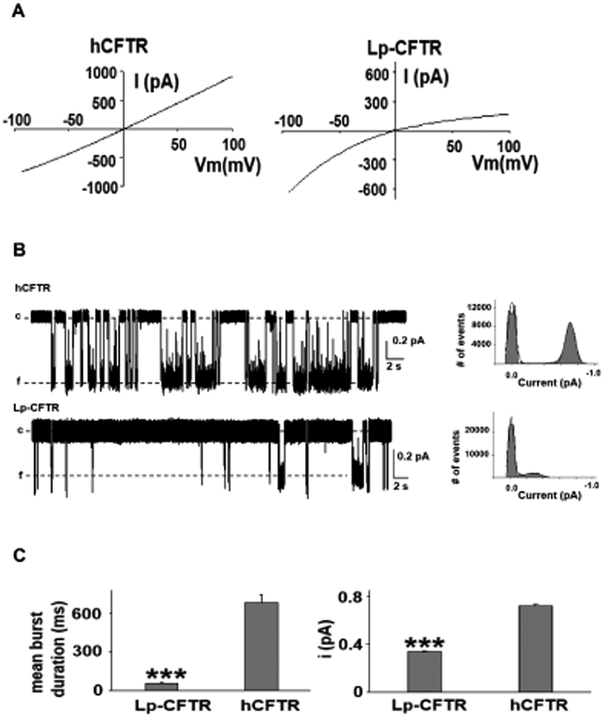
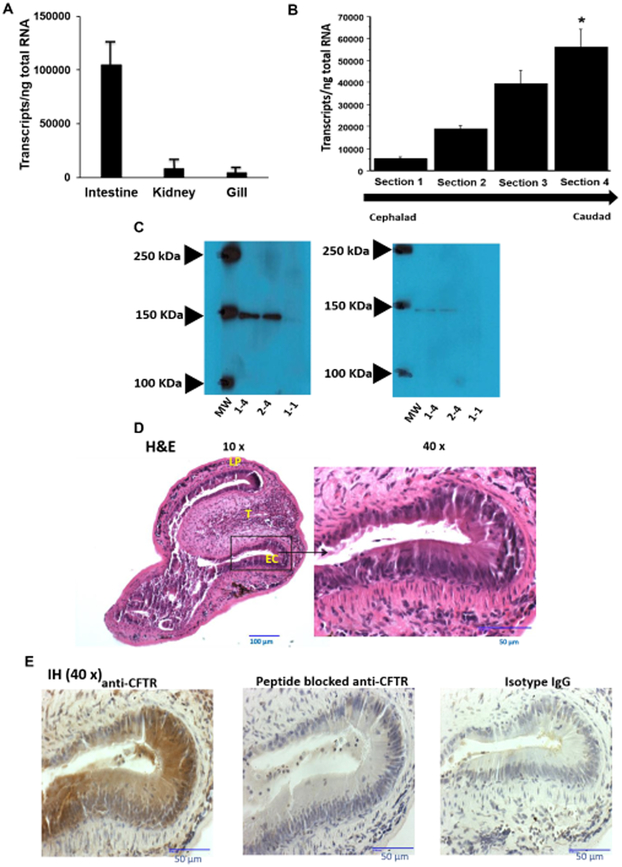
The cystic fibrosis transmembrane conductance regulator (CFTR) is a chloride channel central to the development of secretory diarrhea and cystic fibrosis. The oldest CFTR ortholog identified is from dogfish shark, which retains similar structural and functional characteristics to the mammalian protein, thereby highlighting CFTR's critical role in regulating epithelial ion transport in vertebrates. However, the identification of an early CFTR ortholog with altered structure or function would provide critical insight into the evolution of epithelial anion transport. Here, we describe the earliest known CFTR, expressed in sea lamprey (Petromyzon marinus), with unique structural features, altered kinetics of activation and sensitivity to inhibition, and altered single-channel conductance compared to human CFTR. Our data provide the earliest evolutionary evidence of CFTR, offering insight regarding changes in gene and protein structure that underpin evolution from transporter to anion channel. Importantly, these data provide a unique platform to enhance our understanding of vertebrate phylogeny over a critical period of evolutionary expansion.
Keywords: ABC transporters; CFTR; channel; lamprey; molecular evolution; phosphorylation; vertebrates.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Bessadok A, Garcia E, Jacquet H, Martin S, Garrigues A, Loiseau N, Andre F, Orlowski S, and Vivaudou M (2011). Recognition of sulfonylurea receptor (ABCC8/9) ligands by the multidrug resistance transporter P-glycoprotein (ABCB1): functional similarities based on common structural features between two multispecific ABC proteins. J Biol Chem 286, 3552–3569. - PMC - PubMed
-
- Clemens JD, Nair GB, Ahmed T, Qadri F, and Holmgren J (2017). Cholera. Lancet 390, 1539–1549. - PubMed
-
- Cotten JF, and Welsh MJ (1999). Cystic fibrosis-associated mutations at arginine 347 alter the pore architecture of CFTR. Evidence for disruption of a salt bridge. J Biol Chem 274, 5429–5435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
