

Challenges and Opportunities for Consistent Classification of Human B Cell and Plasma Cell Populations
- PMID: 31681331
- PMCID: PMC6813733
- DOI: 10.3389/fimmu.2019.02458
Challenges and Opportunities for Consistent Classification of Human B Cell and Plasma Cell Populations
Abstract
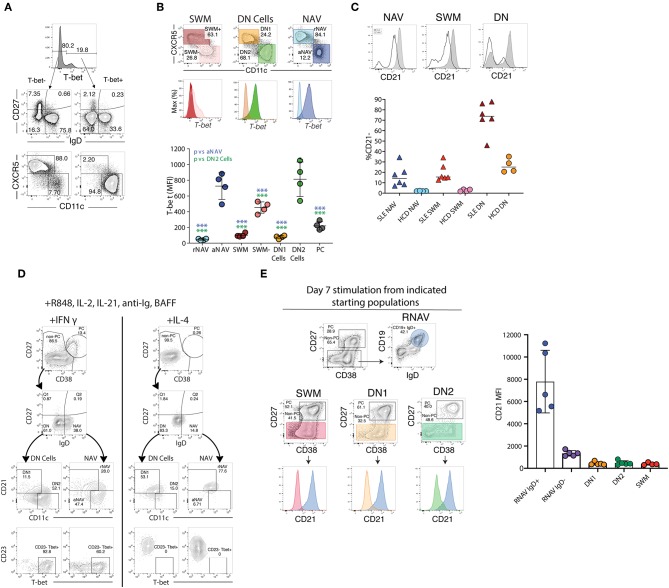
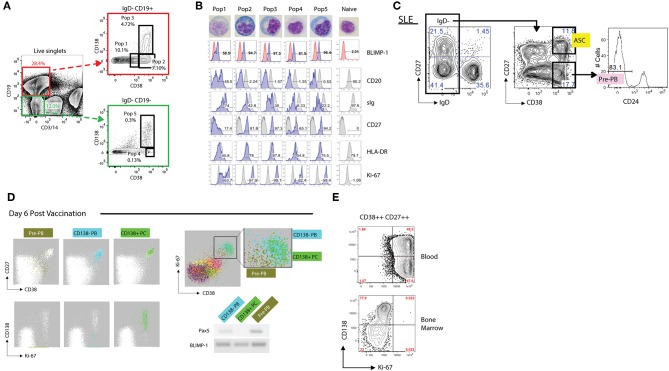
The increasingly recognized role of different types of B cells and plasma cells in protective and pathogenic immune responses combined with technological advances have generated a plethora of information regarding the heterogeneity of this human immune compartment. Unfortunately, the lack of a consistent classification of human B cells also creates significant imprecision on the adjudication of different phenotypes to well-defined populations. Additional confusion in the field stems from: the use of non-discriminatory, overlapping markers to define some populations, the extrapolation of mouse concepts to humans, and the assignation of functional significance to populations often defined by insufficient surface markers. In this review, we shall discuss the current understanding of human B cell heterogeneity and define major parental populations and associated subsets while discussing their functional significance. We shall also identify current challenges and opportunities. It stands to reason that a unified approach will not only permit comparison of separate studies but also improve our ability to define deviations from normative values and to create a clean framework for the identification, functional significance, and disease association with new populations.
Keywords: ABC; B cells; Breg; DN2; atypical B cells; memory; naïve; transitional.
Copyright © 2019 Sanz, Wei, Jenks, Cashman, Tipton, Woodruff, Hom and Lee.
Figures


References
-
- LeBien TW. Fates of human B-cell precursors. Blood. (2000) 96:9–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources