

Range of motion in the avian wing is strongly associated with flight behavior and body mass
- PMID: 31681840
- PMCID: PMC6810231
- DOI: 10.1126/sciadv.aaw6670
Range of motion in the avian wing is strongly associated with flight behavior and body mass
Abstract
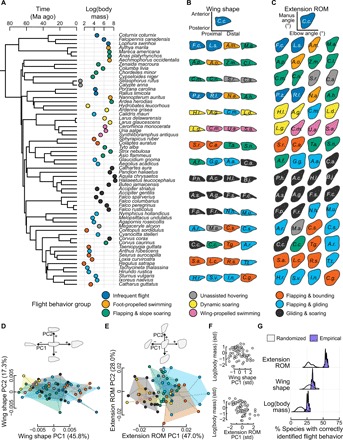
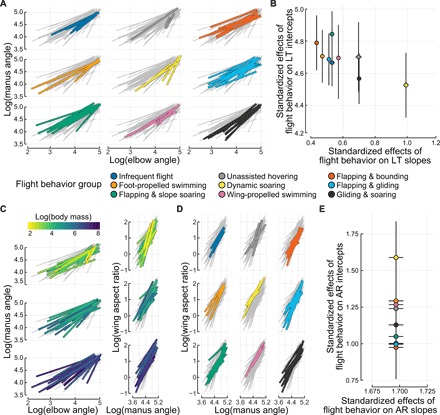
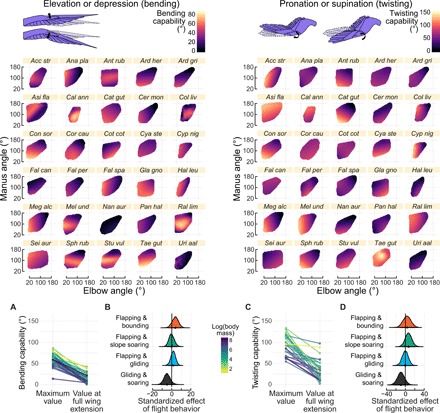
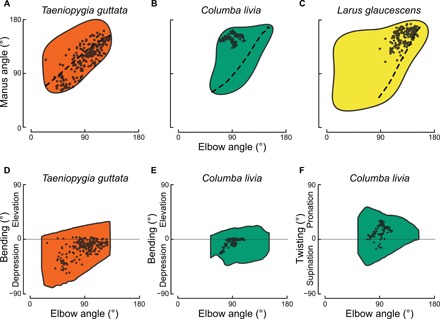
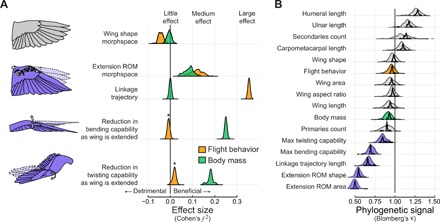
Avian wing shape is highly variable across species but only coarsely associated with flight behavior, performance, and body mass. An underexplored but potentially explanatory feature is the ability of birds to actively change wing shape to meet aerodynamic and behavioral demands. Across 61 species, we found strong associations with flight behavior and mass for range of motion traits but not wing shape and strikingly different associations for different aspects of motion capability. Further, static morphology exhibits high phylogenetic signal, whereas range of motion shows greater evolutionary lability. These results suggest a new framework for understanding the evolution of avian flight: Rather than wing morphology, it is range of motion, an emergent property of morphology, that is predominantly reshaped as flight strategy and body size evolve.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- J. M. V. Rayner, Form and Function in Avian Flight (Current Ornithology series, 1988), vol. 5, pp. 1–66.
-
- Lentink D., Müller U. K., Stamhuis E. J., de Kat R., van Gestel W., Veldhuis L. L., Henningsson P., Hedenström A., Videler J. J., van Leeuwen J. L., How swifts control their glide performance with morphing wings. Nature 446, 1082–1085 (2007). - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
