

Complexes formed with integrin-α5 and KCNB1 potassium channel wild type or epilepsy-susceptibility variants modulate cellular plasticity via Ras and Akt signaling
- PMID: 31682765
- PMCID: PMC6894052
- DOI: 10.1096/fj.201901792R
Complexes formed with integrin-α5 and KCNB1 potassium channel wild type or epilepsy-susceptibility variants modulate cellular plasticity via Ras and Akt signaling
Abstract
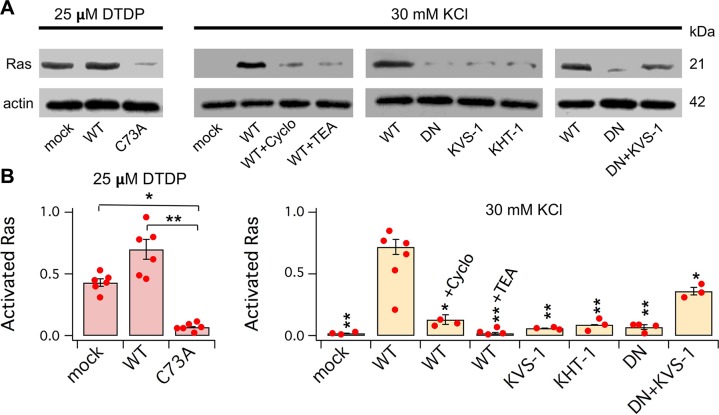
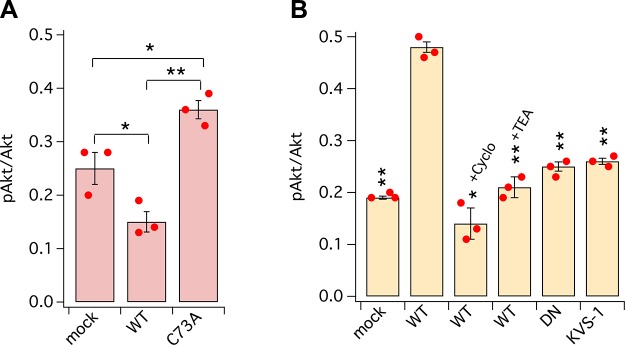
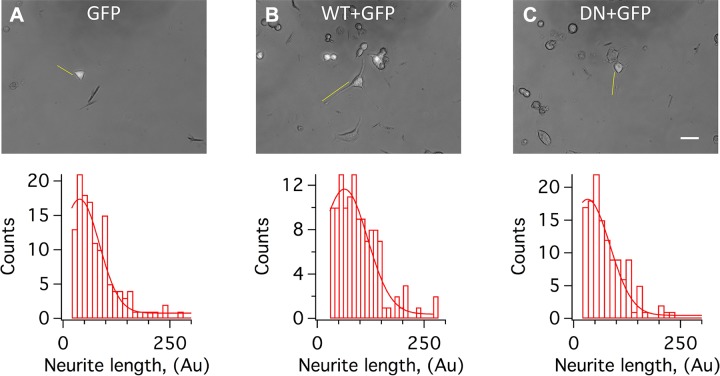
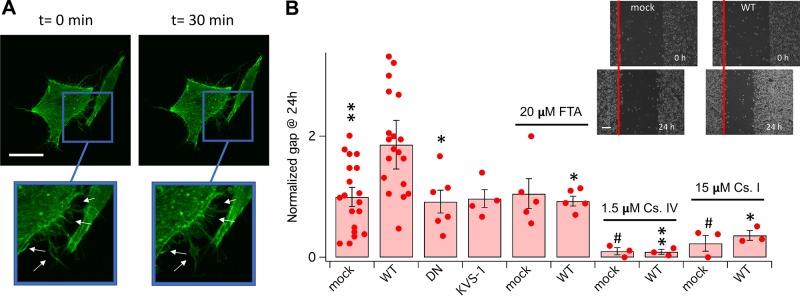
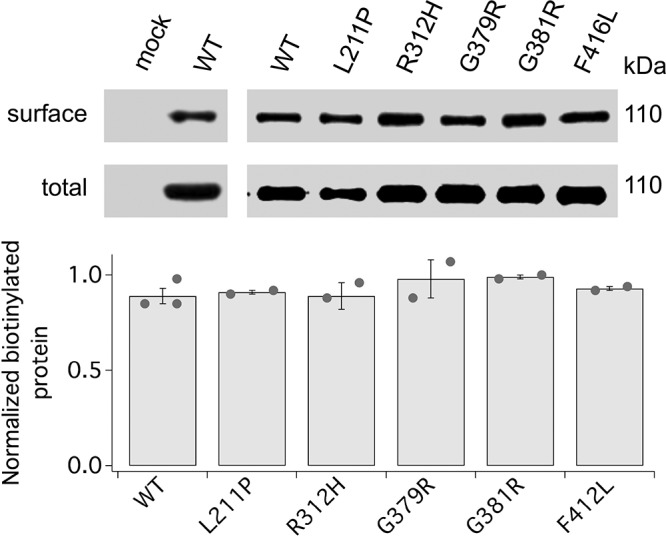
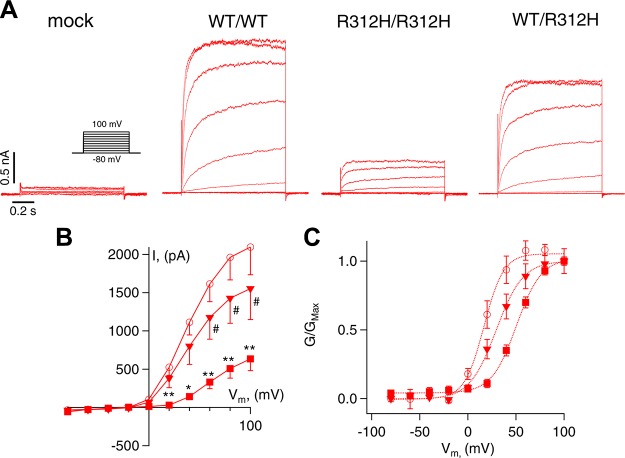
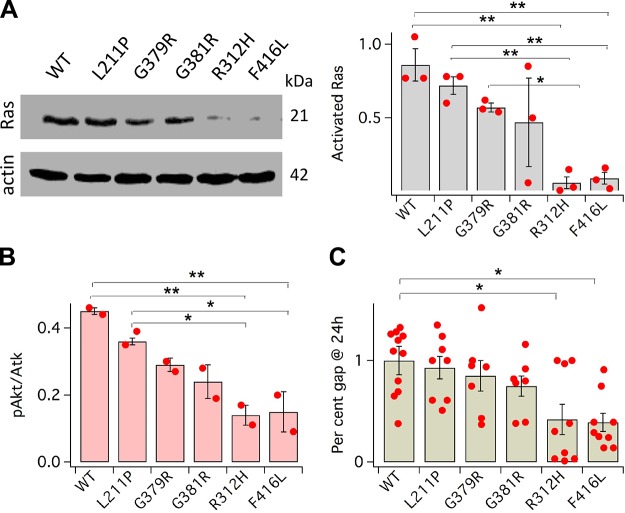
Voltage-gated potassium (K+) channel subfamily B member 1 (KCNB1, Kv2.1) and integrin-α5 form macromolecular complexes-named integrin-α5-KCNB1 complexes (IKCs)-in the human brain, but their function was poorly understood. Here we report that membrane depolarization triggered IKC intracellular signals mediated by small GTPases of the Ras subfamily and protein kinase B (Akt) to advance the development of filopodia and lamellipodia in Chinese hamster ovary cells, stimulate their motility, and enhance neurite outgrowth in mouse neuroblastoma Neuro2a cells. Five KCNB1 mutants (L211P, R312H G379R, G381R, and F416L) linked to severe infancy or early-onset epileptic encephalopathy exhibited markedly defective conduction. However, although L211P, G379R, and G381R normally engaged Ras/Akt and stimulated cell migration, R312H and F416L failed to activate Ras/Akt signaling and did not enhance cell migration. Taken together, these data suggest that IKCs modulate cellular plasticity via Ras and Akt signaling. As such, defective IKCs may cause epilepsy through mechanisms other than dysregulated excitability such as, for example, abnormal neuronal development and resulting synaptic connectivity.-Yu, W., Shin, M. R., Sesti, F. Complexes formed with integrin-α5 and KCNB1 potassium channel wild type or epilepsy-susceptibility variants modulate cellular plasticity via Ras and Akt signaling.
Conflict of interest statement
The authors thank Drs. Mladen-Roko Rasin and Huaye Zhang (Robert Wood Johnson Medical School, Rutgers University) for help with the confocal microscope and Drs. Kiram Madura and Li Chen (Robert Wood Johnson Medical School, Rutgers University) for help with the plate reader. The Lifeact-GFP construct was provided by the Addgene repository. This work was supported by National Science Foundation (NSF) Grant 1456675 and U.S. National Institutes of Health (NIH) Grants R01AG060919 (National Institute on Aging) and R21NS0966 (National Institute of Neurological Disorders and Stroke) to F.S. The authors declare no conflicts of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
