

Genomic analyses of Burkholderia cenocepacia reveal multiple species with differential host-adaptation to plants and humans
- PMID: 31684866
- PMCID: PMC6829993
- DOI: 10.1186/s12864-019-6186-z
Genomic analyses of Burkholderia cenocepacia reveal multiple species with differential host-adaptation to plants and humans
Abstract
Background: Burkholderia cenocepacia is a human opportunistic pathogen causing devastating symptoms in patients suffering from immunodeficiency and cystic fibrosis. Out of the 303 B. cenocepacia strains with available genomes, the large majority were isolated from a clinical context. However, several isolates originate from other environmental sources ranging from aerosols to plant endosphere. Plants can represent reservoirs for human infections as some pathogens can survive and sometimes proliferate in the rhizosphere. We therefore investigated if B. cenocepacia had the same potential.
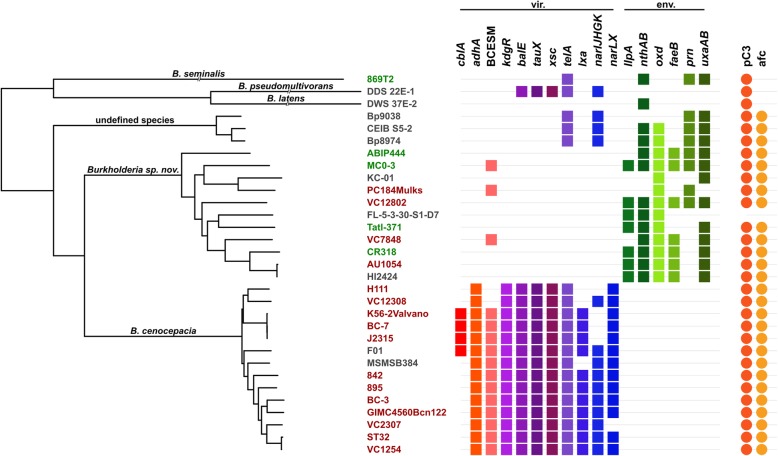
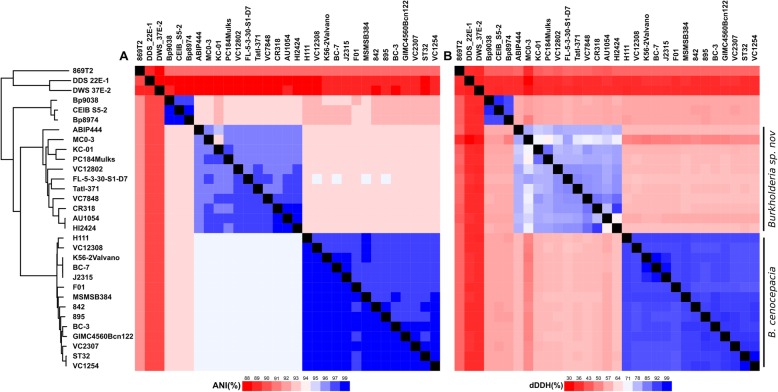
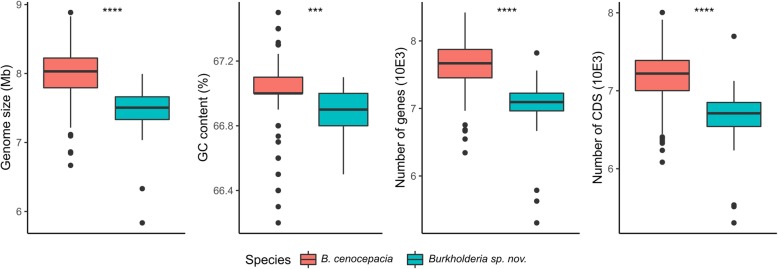
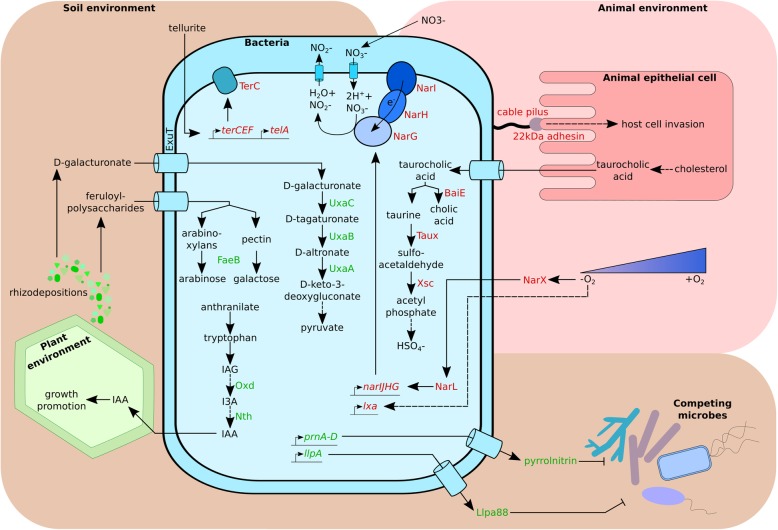
Results: We selected genome sequences from 31 different strains, representative of the diversity of ecological niches of B. cenocepacia, and conducted comparative genomic analyses in the aim of finding specific niche or host-related genetic determinants. Phylogenetic analyses and whole genome average nucleotide identity suggest that strains, registered as B. cenocepacia, belong to at least two different species. Core-genome analyses show that the clade enriched in environmental isolates lacks multiple key virulence factors, which are conserved in the sister clade where most clinical isolates fall, including the highly virulent ET12 lineage. Similarly, several plant associated genes display an opposite distribution between the two clades. Finally, we suggest that B. cenocepacia underwent a host jump from plants/environment to animals, as supported by the phylogenetic analysis. We eventually propose a name for the new species that lacks several genetic traits involved in human virulence.
Conclusion: Regardless of the method used, our studies resulted in a disunited perspective of the B. cenocepacia species. Strains currently affiliated to this taxon belong to at least two distinct species, one having lost several determining animal virulence factors.
Keywords: Burkholderia cenocepacia; Comparative genomics; Host adaptation; Opportunistic pathogen; PGPR.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Estrada-de los Santos P, Palmer M, Chávez-Ramírez B, Beukes C, Steenkamp E, Briscoe L, et al. Whole genome analyses suggests that burkholderia sensu lato contains two additional novel genera (mycetohabitans gen. nov., and trinickia gen. nov.): implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes (Basel) 2018;9:389. doi: 10.3390/genes9080389. - DOI - PMC - PubMed
-
- Dobritsa AP, Samadpour M. Transfer of eleven species of the genus Burkholderia to the genus Paraburkholderia and proposal of Caballeronia gen. Nov. to accommodate twelve species of the genera Burkholderia and Paraburkholderia. Int J Syst Evol Microbiol. 2016;66:2836–2846. doi: 10.1099/ijsem.0.001065. - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
