Developmental Plasticity and Cellular Reprogramming in Caenorhabditis elegans
- PMID: 31685551
- PMCID: PMC6827377
- DOI: 10.1534/genetics.119.302333
Developmental Plasticity and Cellular Reprogramming in Caenorhabditis elegans
Abstract
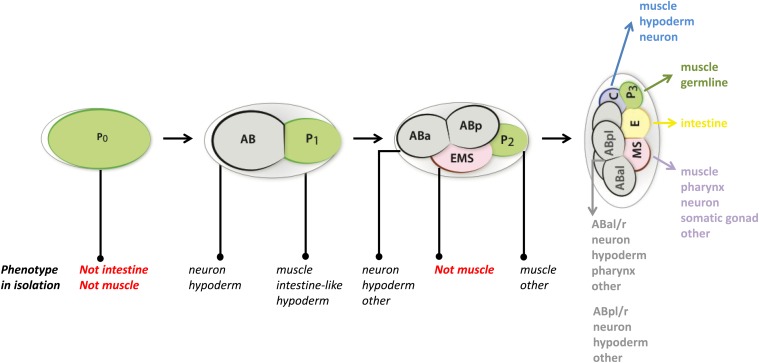
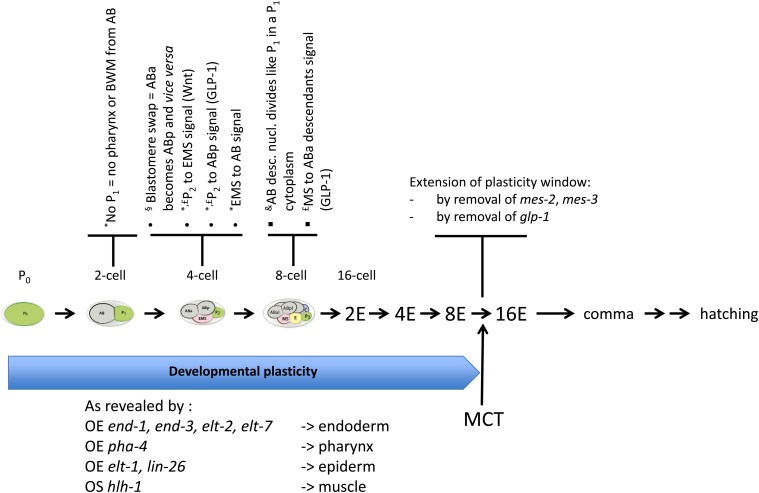
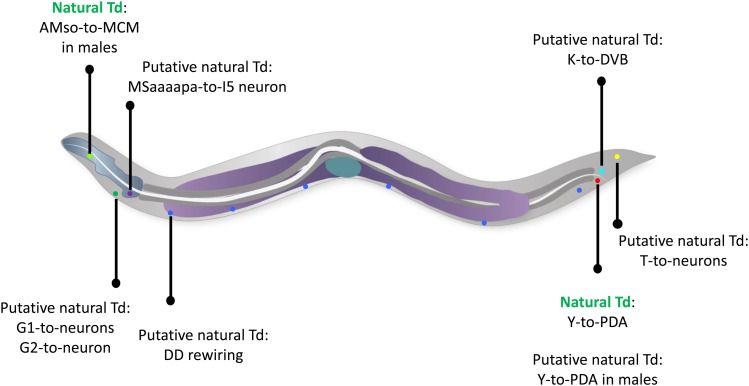
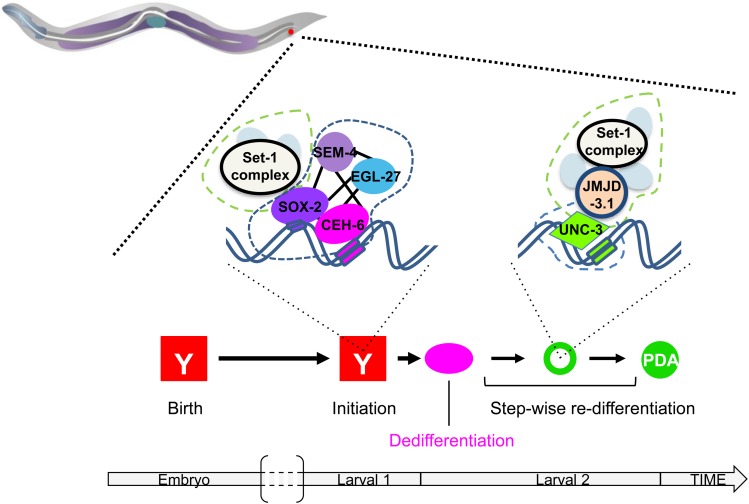
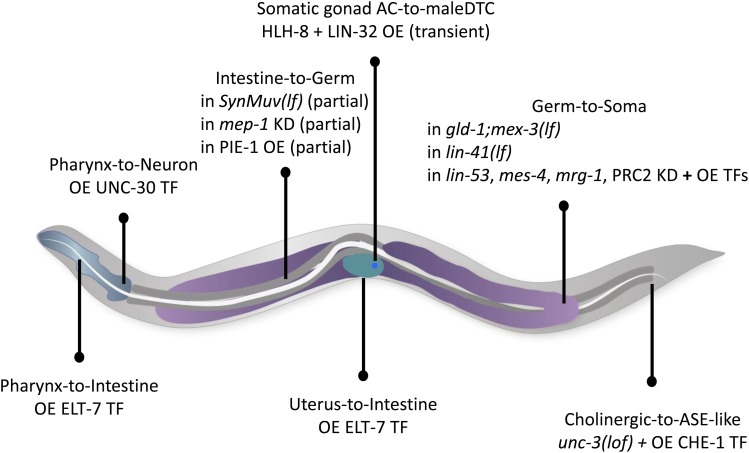
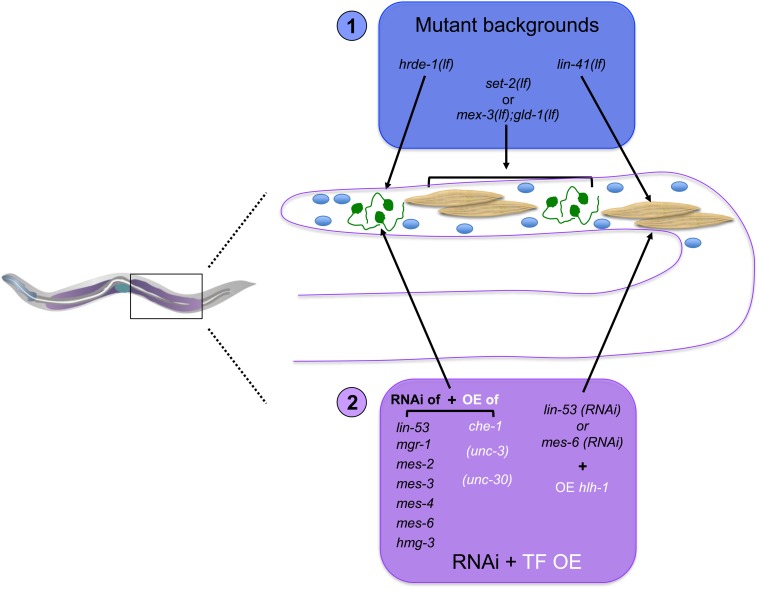
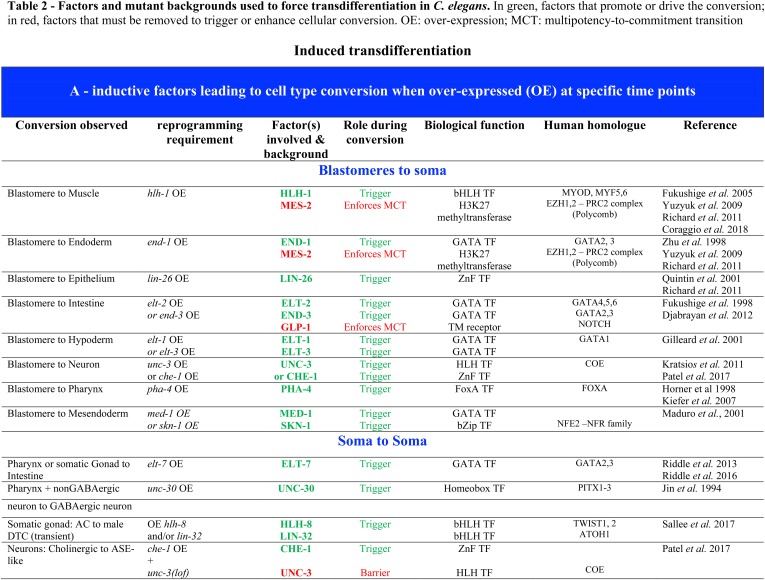
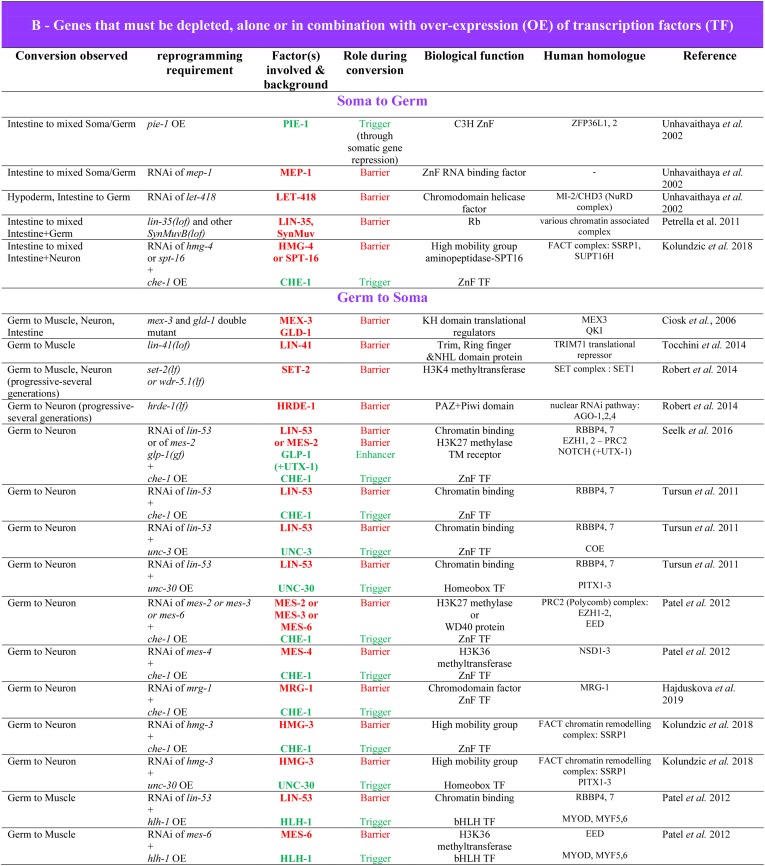
While Caenorhabditis elegans was originally regarded as a model for investigating determinate developmental programs, landmark studies have subsequently shown that the largely invariant pattern of development in the animal does not reflect irreversibility in rigidly fixed cell fates. Rather, cells at all stages of development, in both the soma and germline, have been shown to be capable of changing their fates through mutation or forced expression of fate-determining factors, as well as during the normal course of development. In this chapter, we review the basis for natural and induced cellular plasticity in C. elegans We describe the events that progressively restrict cellular differentiation during embryogenesis, starting with the multipotency-to-commitment transition (MCT) and subsequently through postembryonic development of the animal, and consider the range of molecular processes, including transcriptional and translational control systems, that contribute to cellular plasticity. These findings in the worm are discussed in the context of both classical and recent studies of cellular plasticity in vertebrate systems.
Keywords: WormBook; cell type conversion; reprogramming; stem cells; transdetermination; transdifferentiation.
Copyright © 2019 by the Genetics Society of America.
Figures