

A chromosome folding intermediate at the condensin-to-cohesin transition during telophase
- PMID: 31685986
- PMCID: PMC6858582
- DOI: 10.1038/s41556-019-0406-2
A chromosome folding intermediate at the condensin-to-cohesin transition during telophase
Abstract
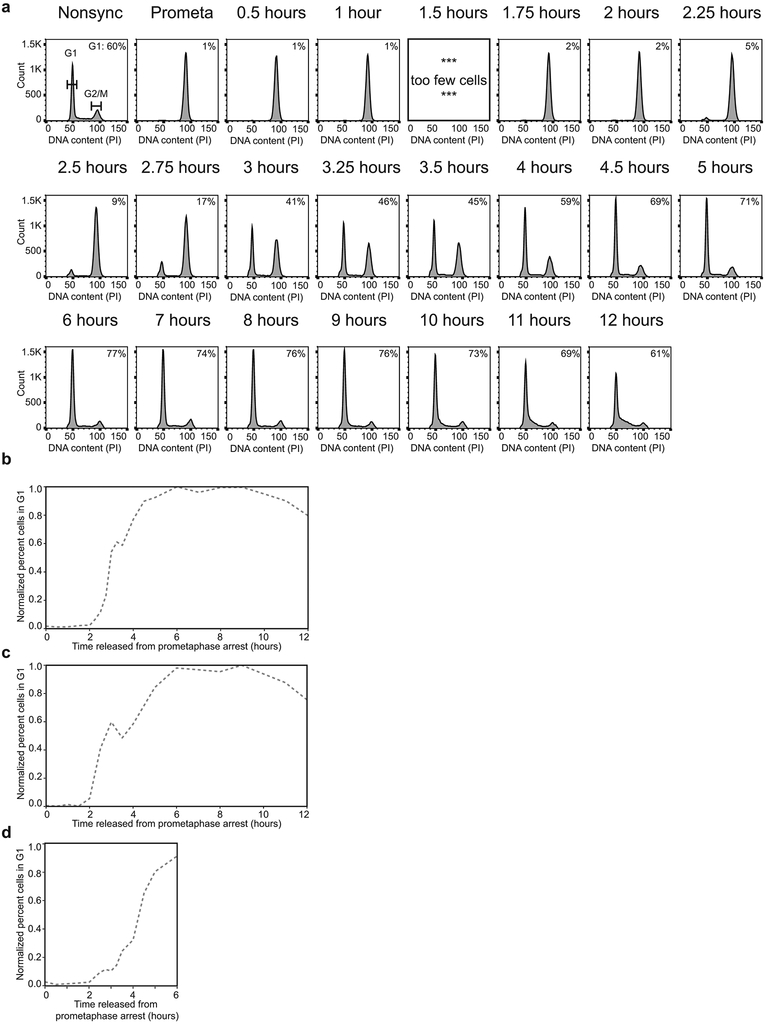
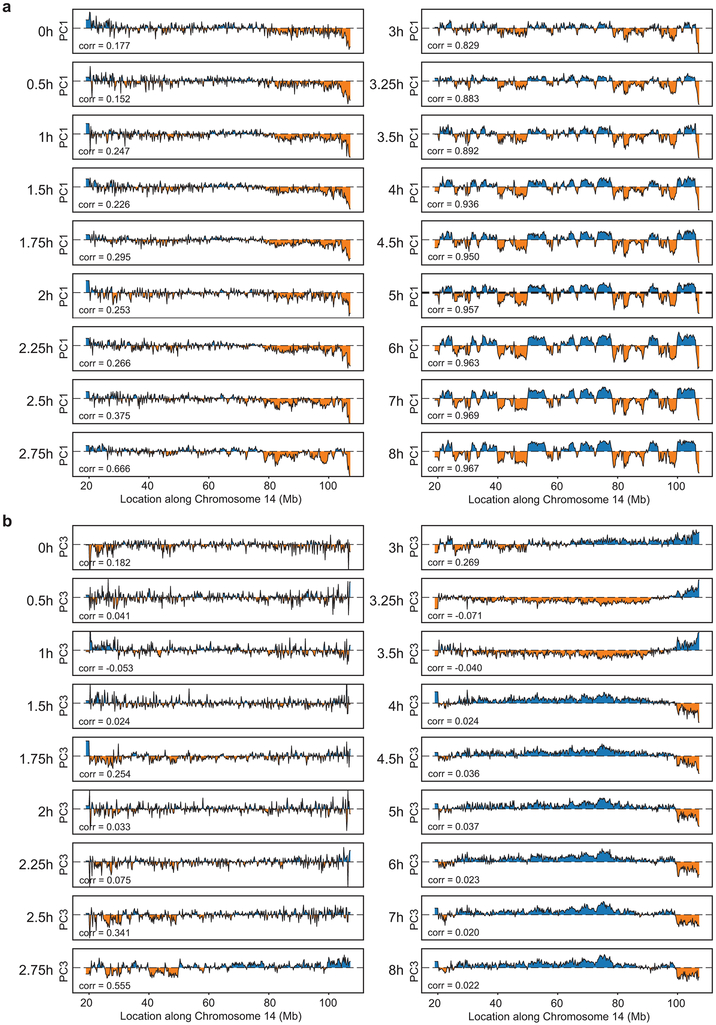
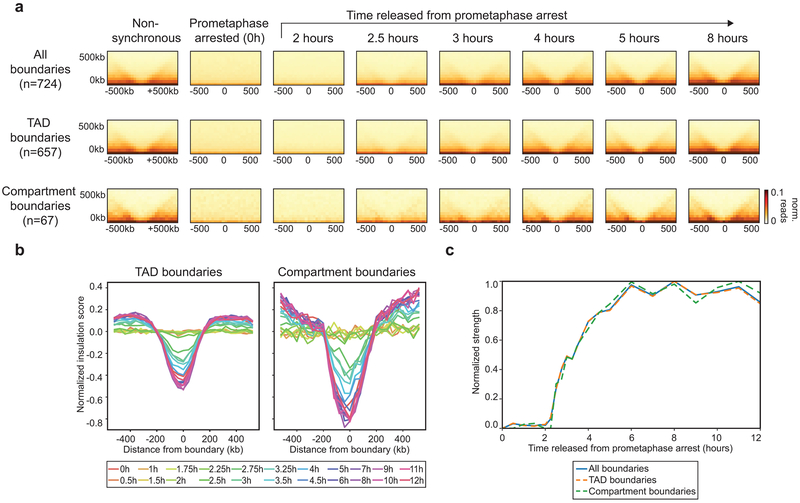
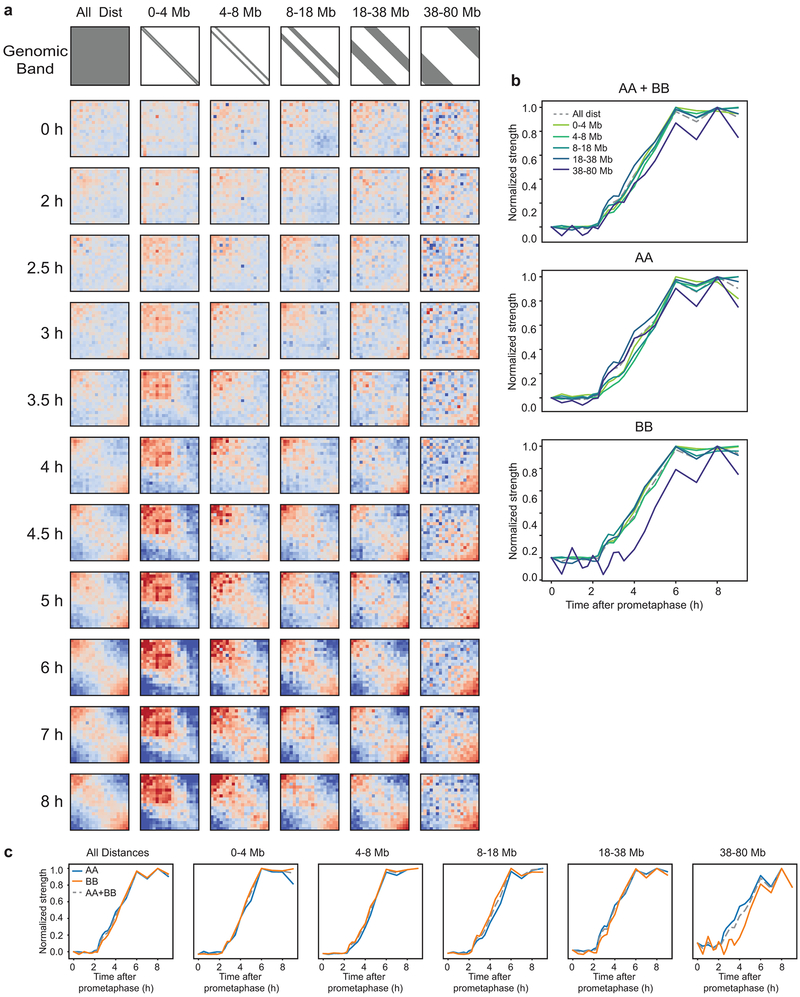
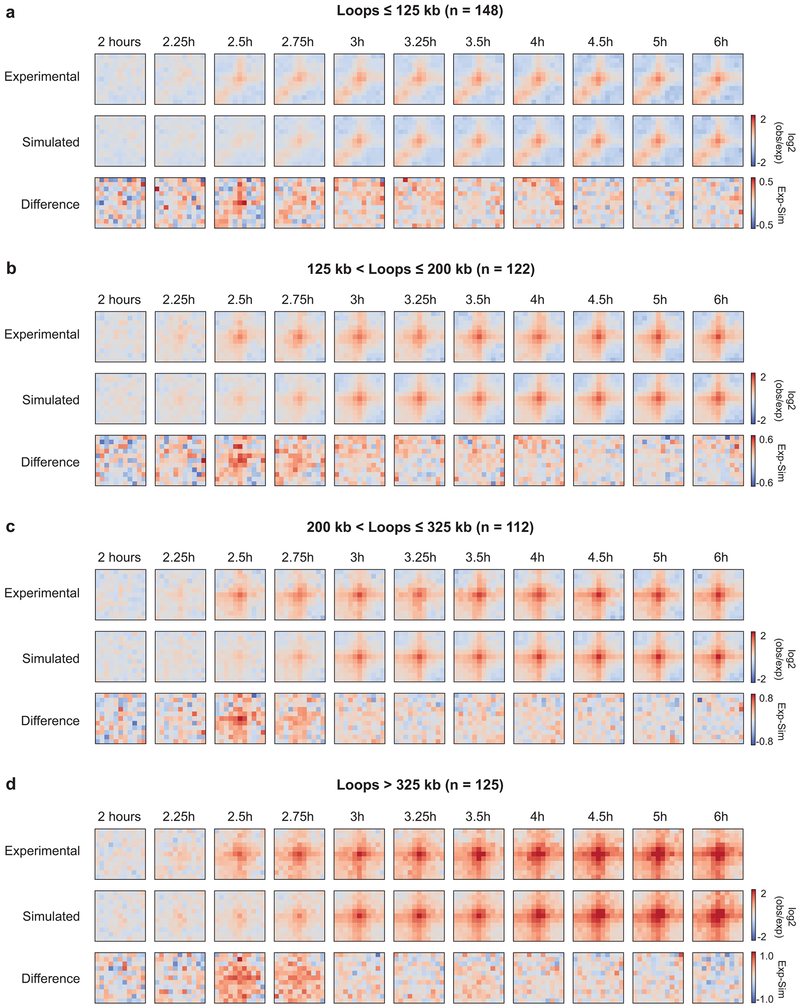
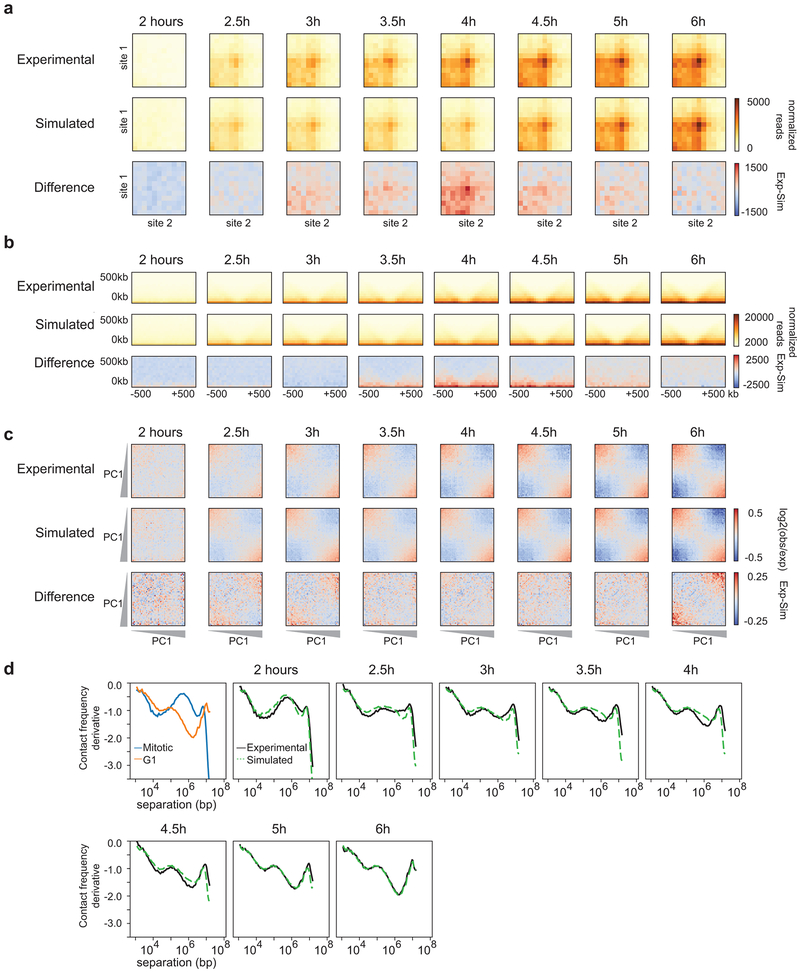
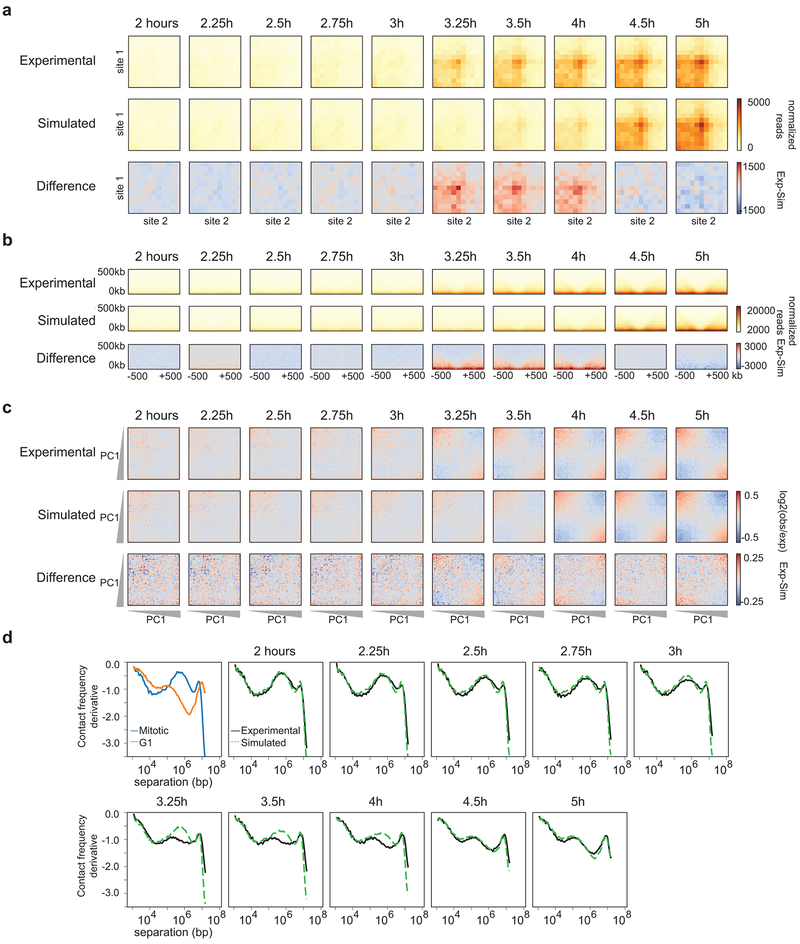
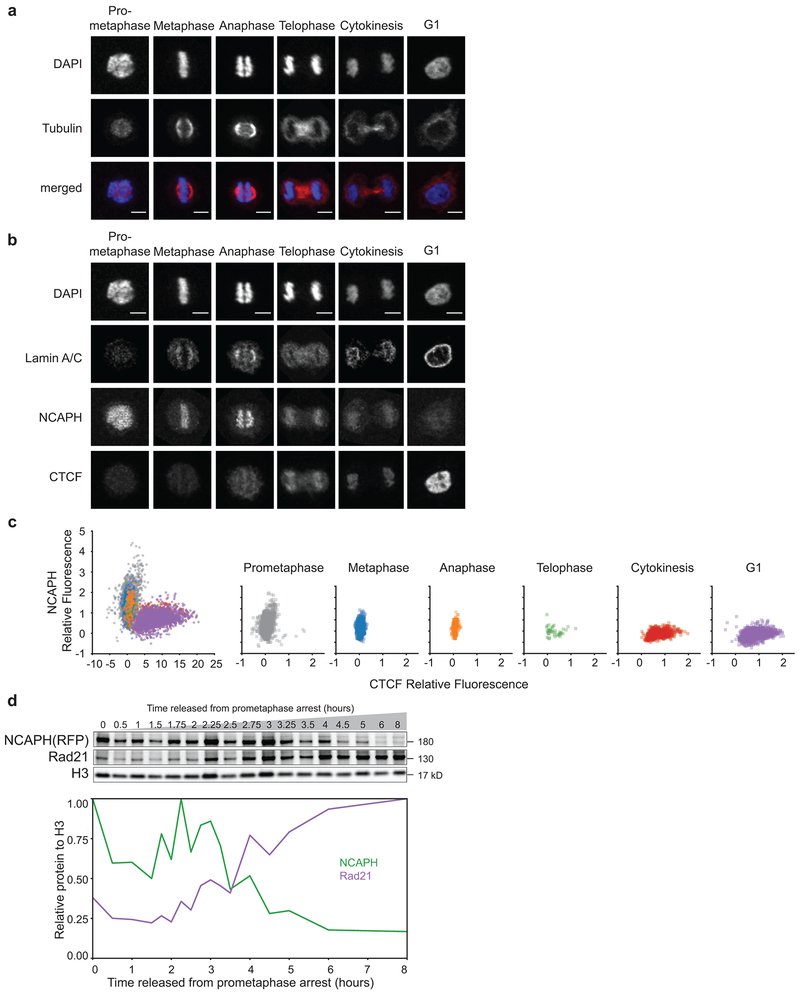
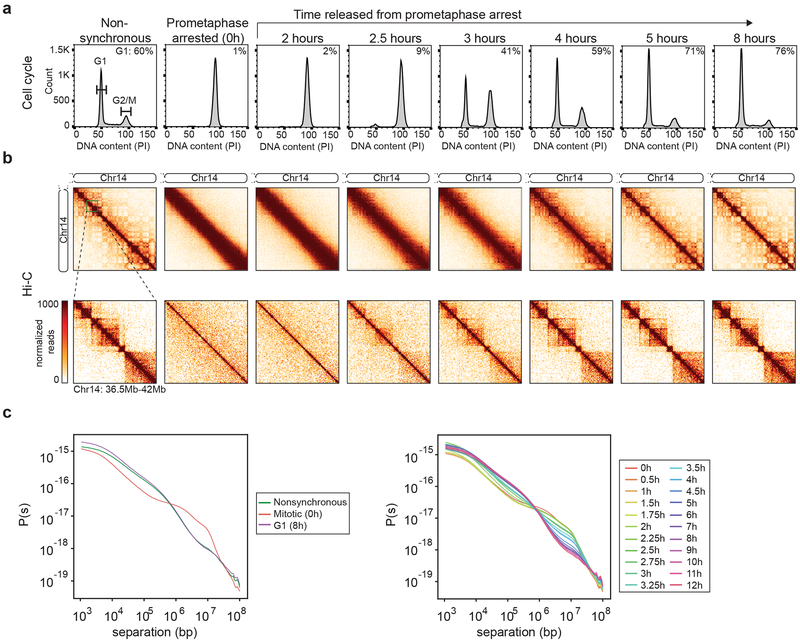
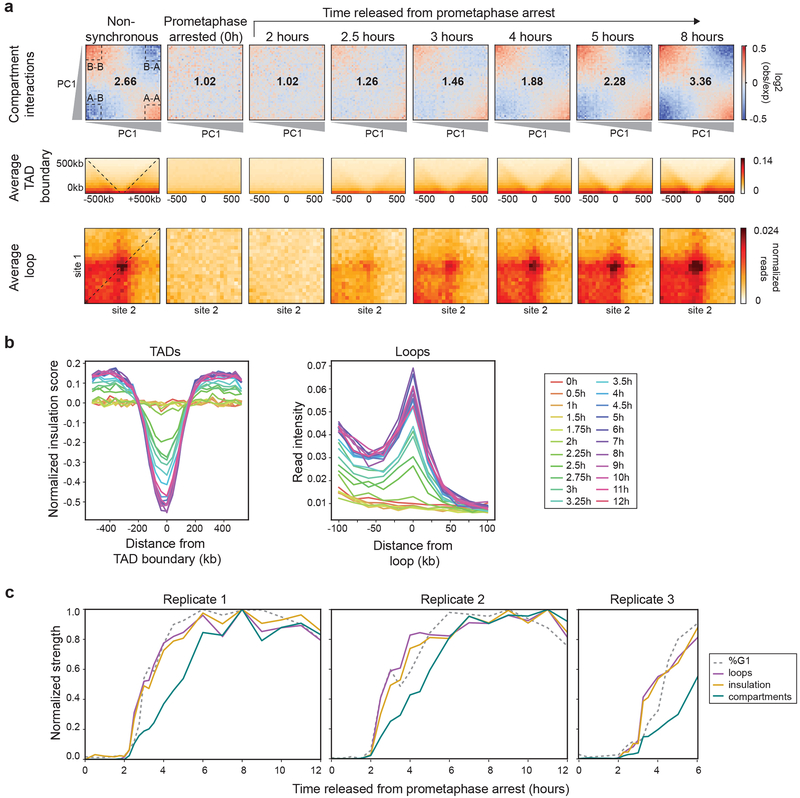
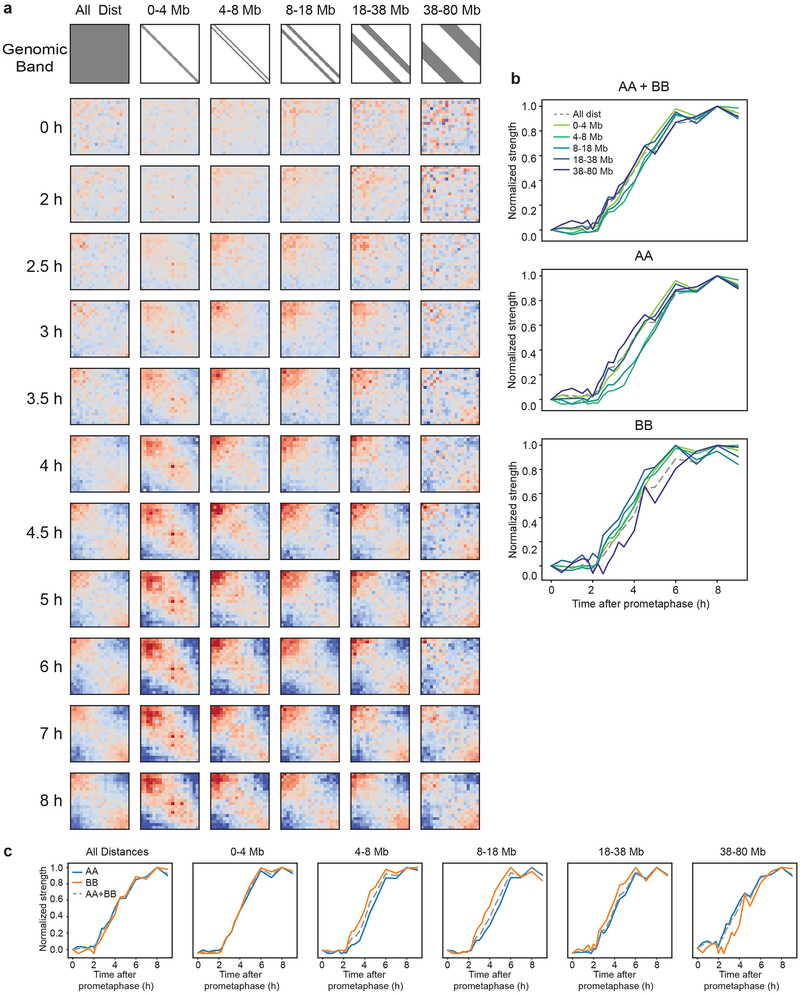
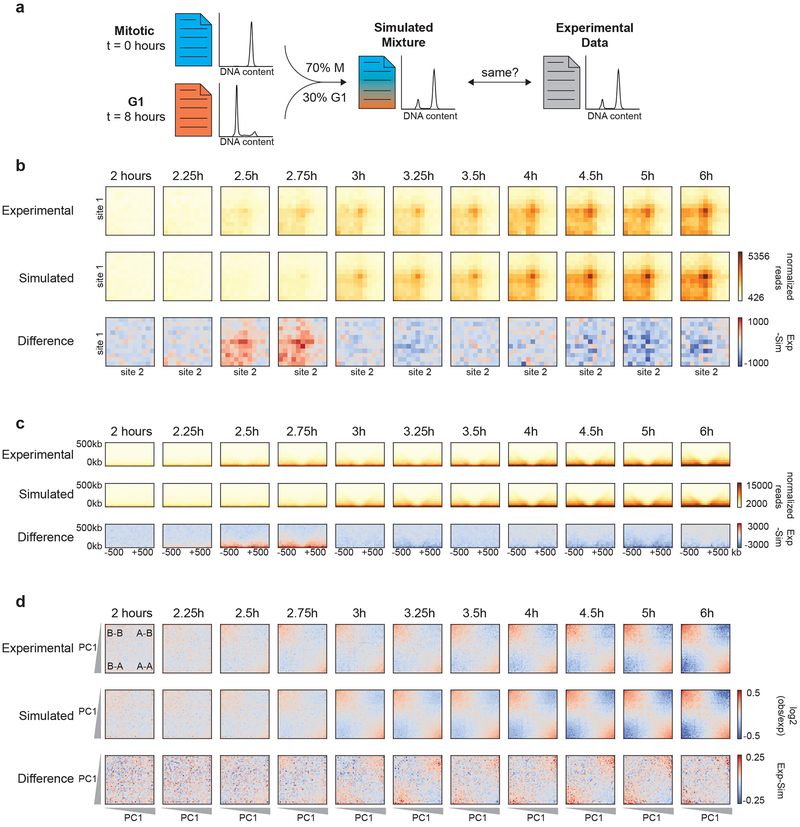
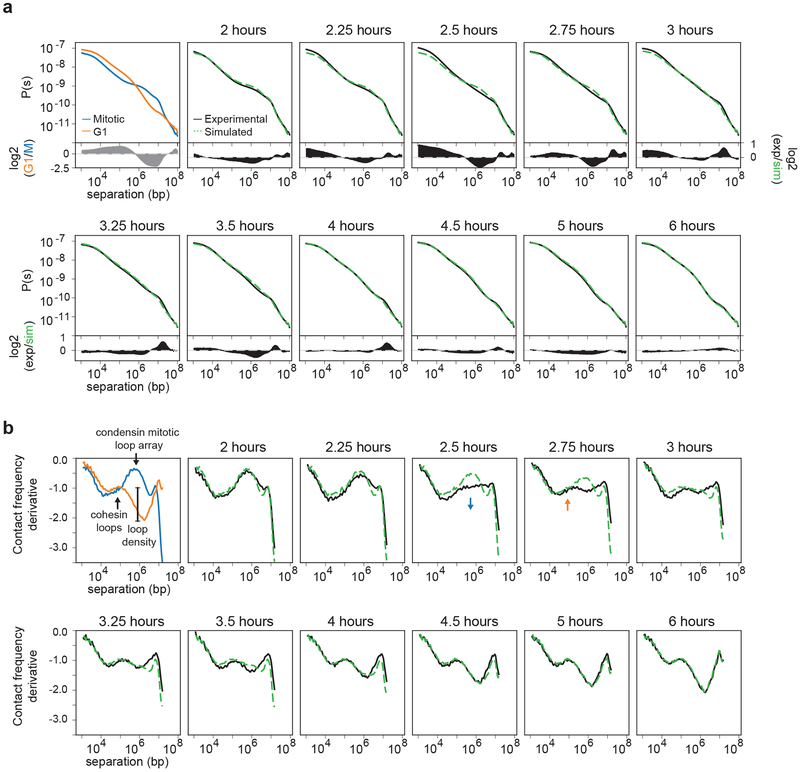
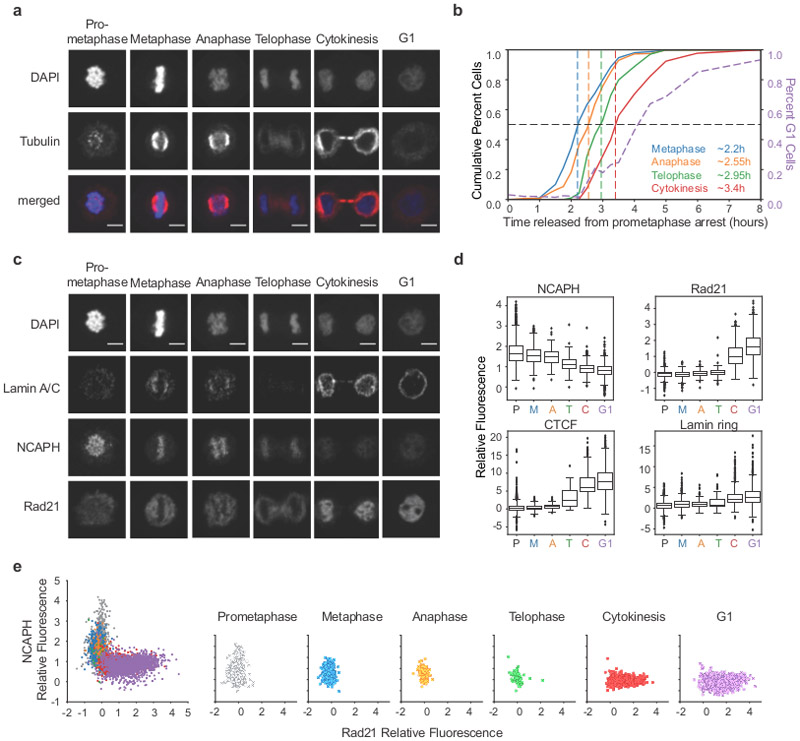
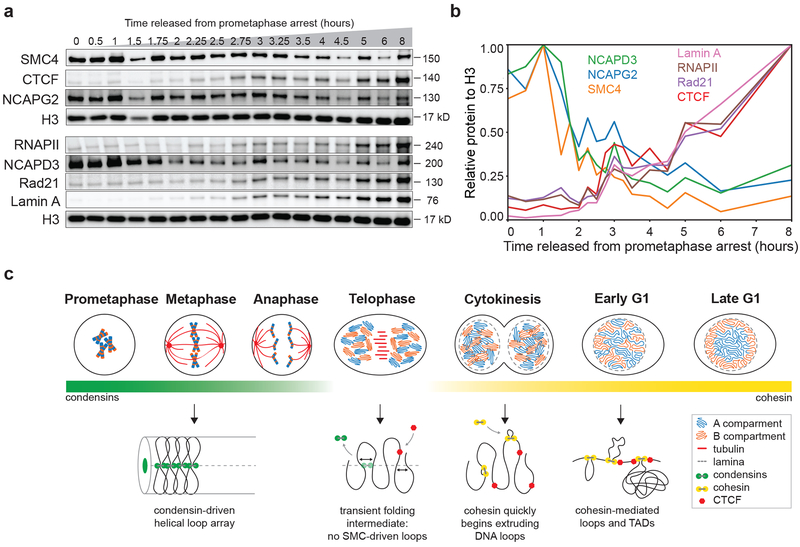
Chromosome folding is modulated as cells progress through the cell cycle. During mitosis, condensins fold chromosomes into helical loop arrays. In interphase, the cohesin complex generates loops and topologically associating domains (TADs), while a separate process of compartmentalization drives segregation of active and inactive chromatin. We used synchronized cell cultures to determine how the mitotic chromosome conformation transforms into the interphase state. Using high-throughput chromosome conformation capture (Hi-C) analysis, chromatin binding assays and immunofluorescence, we show that, by telophase, condensin-mediated loops are lost and a transient folding intermediate is formed that is devoid of most loops. By cytokinesis, cohesin-mediated CTCF-CTCF loops and the positions of TADs emerge. Compartment boundaries are also established early, but long-range compartmentalization is a slow process and proceeds for hours after cells enter G1. Our results reveal the kinetics and order of events by which the interphase chromosome state is formed and identify telophase as a critical transition between condensin- and cohesin-driven chromosome folding.
Figures
Comment in
-
A transient absence of SMC-mediated loops after mitosis.Nat Cell Biol. 2019 Nov;21(11):1303-1304. doi: 10.1038/s41556-019-0421-3. Nat Cell Biol. 2019. PMID: 31685985 No abstract available.
References
-
- de Wit E et al. CTCF Binding Polarity Determines Chromatin Looping. Mol Cell. 60, 676–684 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
