

The molecular architecture of photoreceptor phosphodiesterase 6 (PDE6) with activated G protein elucidates the mechanism of visual excitation
- PMID: 31690623
- PMCID: PMC6926442
- DOI: 10.1074/jbc.RA119.011002
The molecular architecture of photoreceptor phosphodiesterase 6 (PDE6) with activated G protein elucidates the mechanism of visual excitation
Abstract
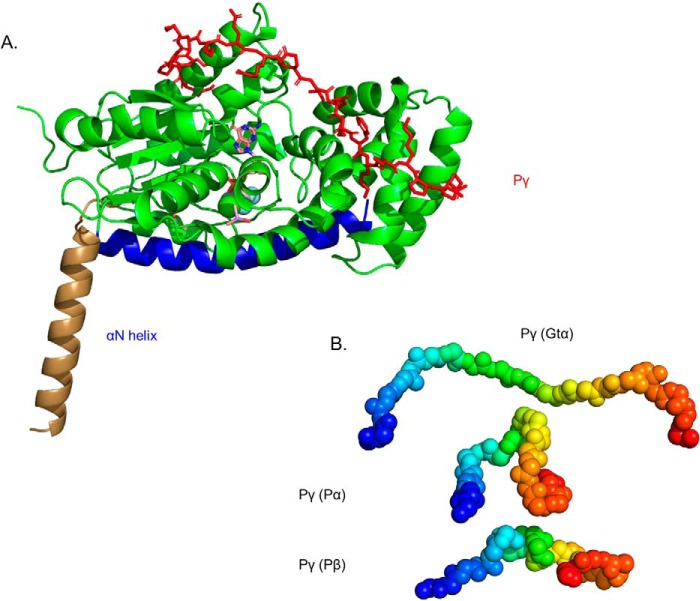
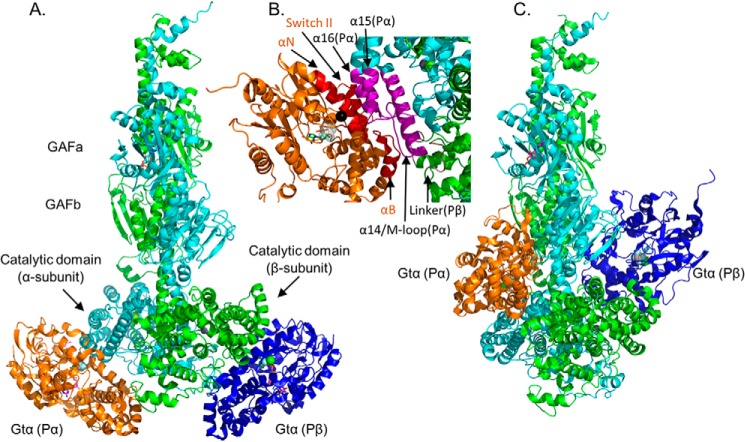
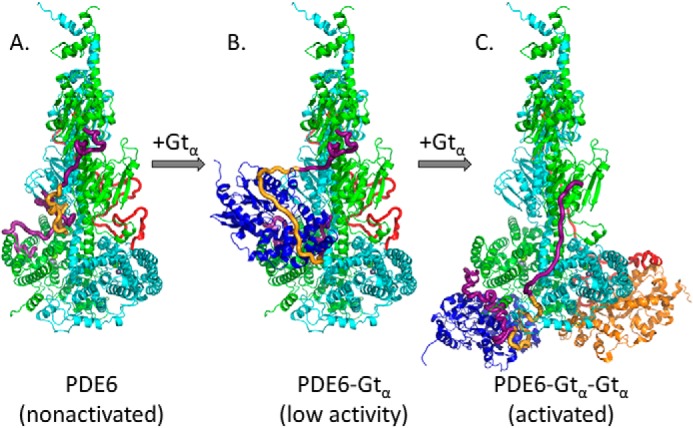
Photoreceptor phosphodiesterase 6 (PDE6) is the central effector of the visual excitation pathway in both rod and cone photoreceptors, and PDE6 mutations that alter PDE6 structure or regulation can result in several human retinal diseases. The rod PDE6 holoenzyme consists of two catalytic subunits (Pαβ) whose activity is suppressed in the dark by binding of two inhibitory γ-subunits (Pγ). Upon photoactivation of rhodopsin, the heterotrimeric G protein (transducin) is activated, resulting in binding of the activated transducin α-subunit (Gtα) to PDE6, displacement of Pγ from the PDE6 active site, and enzyme activation. Although the biochemistry of this pathway is understood, a lack of detailed structural information about the PDE6 activation mechanism hampers efforts to develop therapeutic interventions for managing PDE6-associated retinal diseases. To address this gap, here we used a cross-linking MS-based approach to create a model of the entire interaction surface of Pγ with the regulatory and catalytic domains of Pαβ in its nonactivated state. Following reconstitution of PDE6 and activated Gtα with liposomes and identification of cross-links between Gtα and PDE6 subunits, we determined that the PDE6-Gtα protein complex consists of two Gtα-binding sites per holoenzyme. Each Gtα interacts with the catalytic domains of both catalytic subunits and induces major changes in the interaction sites of the Pγ subunit with the catalytic subunits. These results provide the first structural model for the activated state of the transducin-PDE6 complex during visual excitation, enhancing our understanding of the molecular etiology of inherited retinal diseases.
Keywords: G protein; PDE6; allosteric regulation; integrative structural modeling; mass spectrometry (MS); phosphodiesterases; photoreceptor; phototransduction; protein cross-linking; transducin.
© 2019 Irwin et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

References
-
- Cote R. H. (2006) Photoreceptor phosphodiesterase (PDE6): a G-protein-activated PDE regulating visual excitation in rod and cone photoreceptor cells. In Cyclic Nucleotide Phosphodiesterases in Health and Disease (Beavo J. A., Francis S. H., and Houslay M. D., eds) pp. 165–193, CRC Press, Boca Raton, FL
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
