

How fly neurons compute the direction of visual motion
- PMID: 31691093
- PMCID: PMC7069908
- DOI: 10.1007/s00359-019-01375-9
How fly neurons compute the direction of visual motion
Abstract
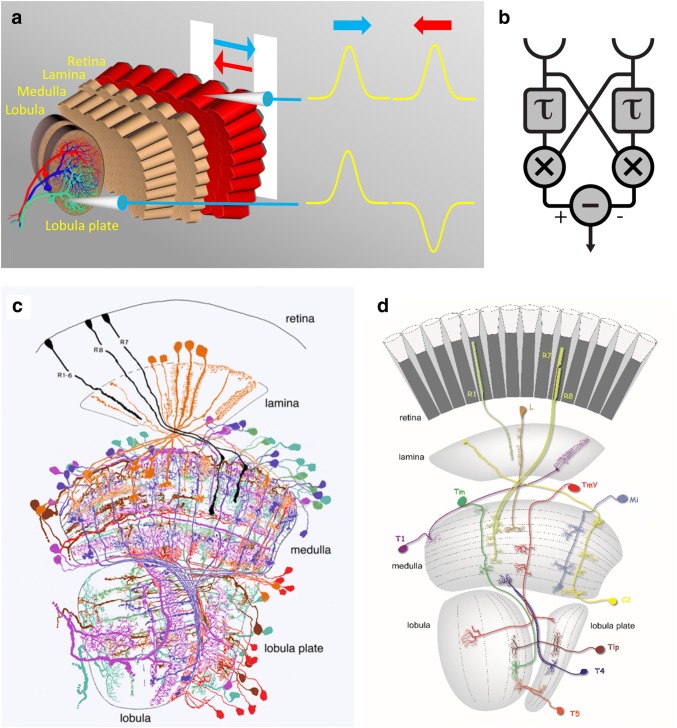
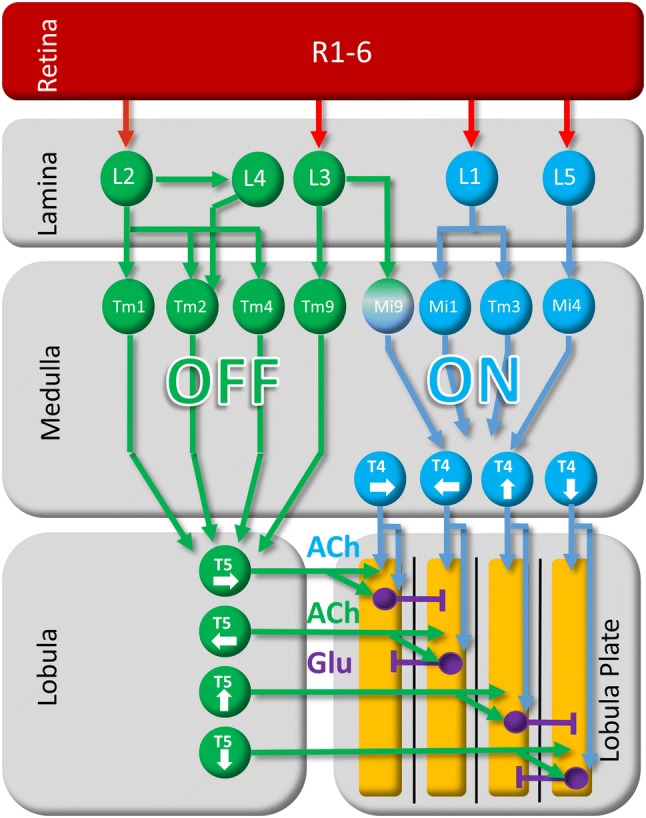
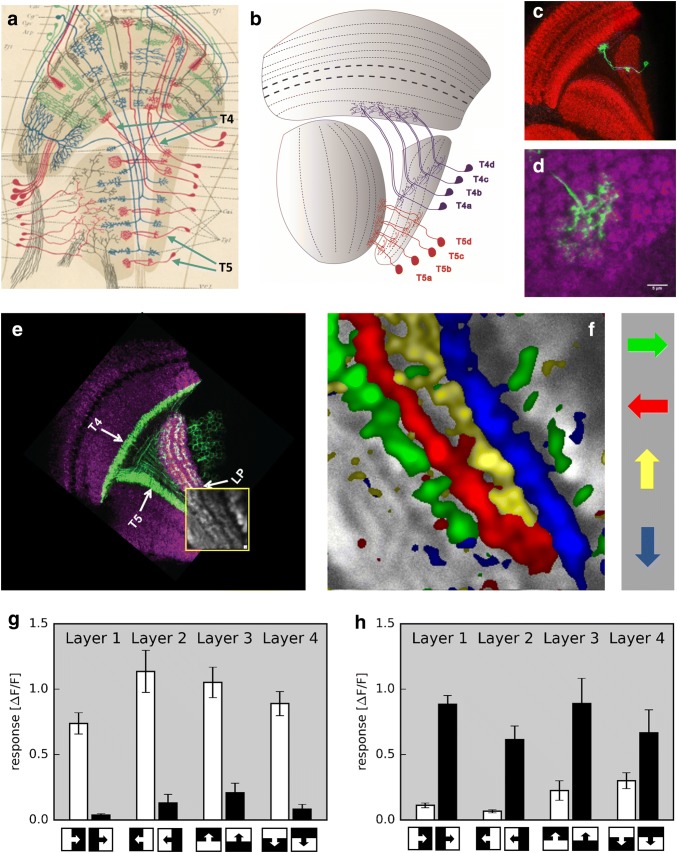
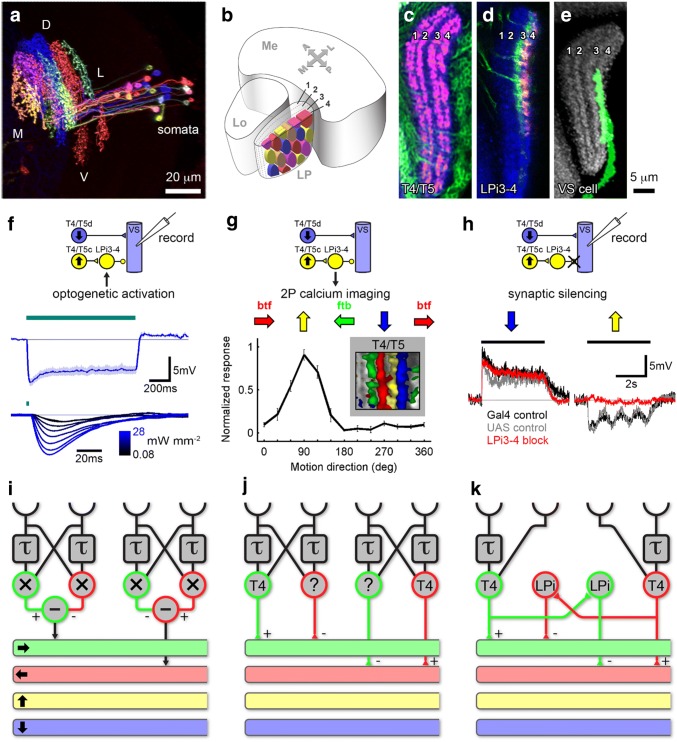
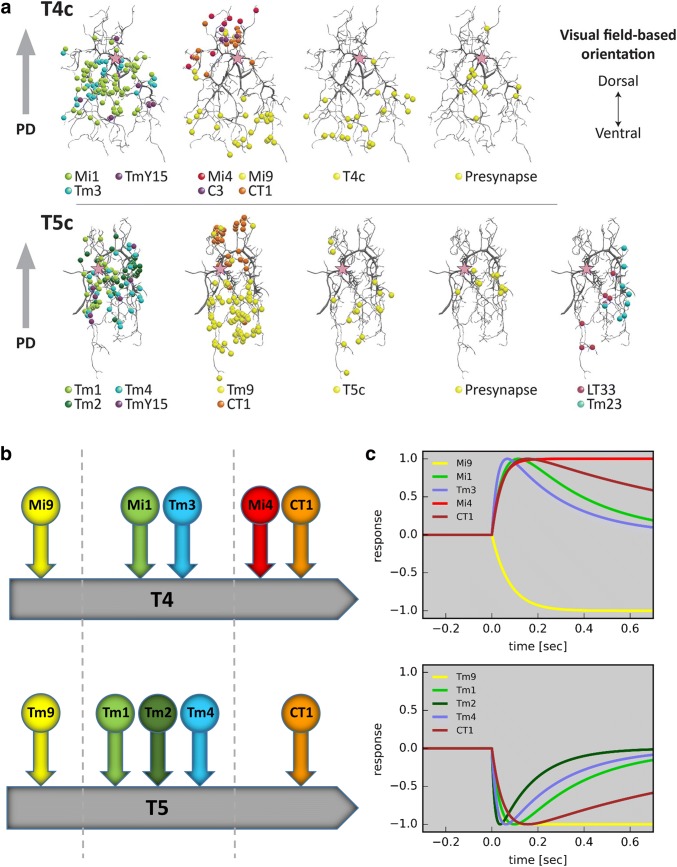
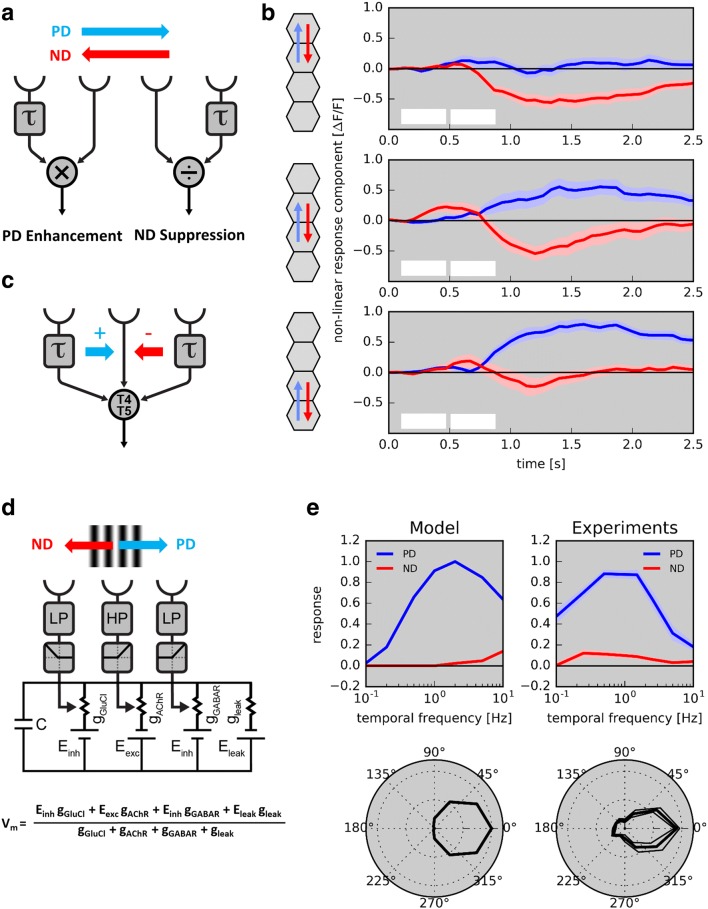
Detecting the direction of image motion is a fundamental component of visual computation, essential for survival of the animal. However, at the level of individual photoreceptors, the direction in which the image is shifting is not explicitly represented. Rather, directional motion information needs to be extracted from the photoreceptor array by comparing the signals of neighboring units over time. The exact nature of this process as implemented in the visual system of the fruit fly Drosophila melanogaster has been studied in great detail, and much progress has recently been made in determining the neural circuits giving rise to directional motion information. The results reveal the following: (1) motion information is computed in parallel ON and OFF pathways. (2) Within each pathway, T4 (ON) and T5 (OFF) cells are the first neurons to represent the direction of motion. Four subtypes of T4 and T5 cells exist, each sensitive to one of the four cardinal directions. (3) The core process of direction selectivity as implemented on the dendrites of T4 and T5 cells comprises both an enhancement of signals for motion along their preferred direction as well as a suppression of signals for motion along the opposite direction. This combined strategy ensures a high degree of direction selectivity right at the first stage where the direction of motion is computed. (4) At the subsequent processing stage, tangential cells spatially integrate direct excitation from ON and OFF-selective T4 and T5 cells and indirect inhibition from bi-stratified LPi cells activated by neighboring T4/T5 terminals, thus generating flow-field-selective responses.
Keywords: Direction selectivity; Drosophila; Null direction suppression; Optic lobe; Preferred direction enhancement; Visual motion.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Akerboom J, Chen TW, Wardill TJ, Tian L, Marvin JS, Mutlu S, Calderón NC, Esposti F, Borghuis BG, Sun XR, Gordus A, Orger MB, Portugues R, Engert F, Macklin JJ, Filosa A, Aggarwal A, Kerr RA, Takagi R, Kracun S, Shigetomi E, Khakh BS, Baier H, Lagnado L, Wang SS, Bargmann CI, Kimmel BE, Jayaraman V, Svoboda K, Kim DS, Schreiter ER, Looger LL. Optimization of a GCaMP calcium indicator for neural activity imaging. J Neurosci. 2012;32:13819–13840. - PMC - PubMed
-
- Ammer G, Leonhardt A, Bahl A, Dickson BJ, Borst A. Functional specialization of neural input elements to the Drosophila ON motion detector. Curr Biol. 2015;25:2247–2253. - PubMed
-
- Anstis SM, Rogers BJ. Illusory reversal of visual depth and movement during changes of contrast. Vision Res. 1975;15:957–961. - PubMed
-
- Arenz A, Drews MS, Richter FG, Ammer G, Borst A. The temporal tuning of the Drosophila motion detectors is determined by the dynamics of their input elements. Curr Biol. 2017;27:929–944. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
