

Changes to the TDP-43 and FUS Interactomes Induced by DNA Damage
- PMID: 31693373
- PMCID: PMC6947635
- DOI: 10.1021/acs.jproteome.9b00575
Changes to the TDP-43 and FUS Interactomes Induced by DNA Damage
Abstract
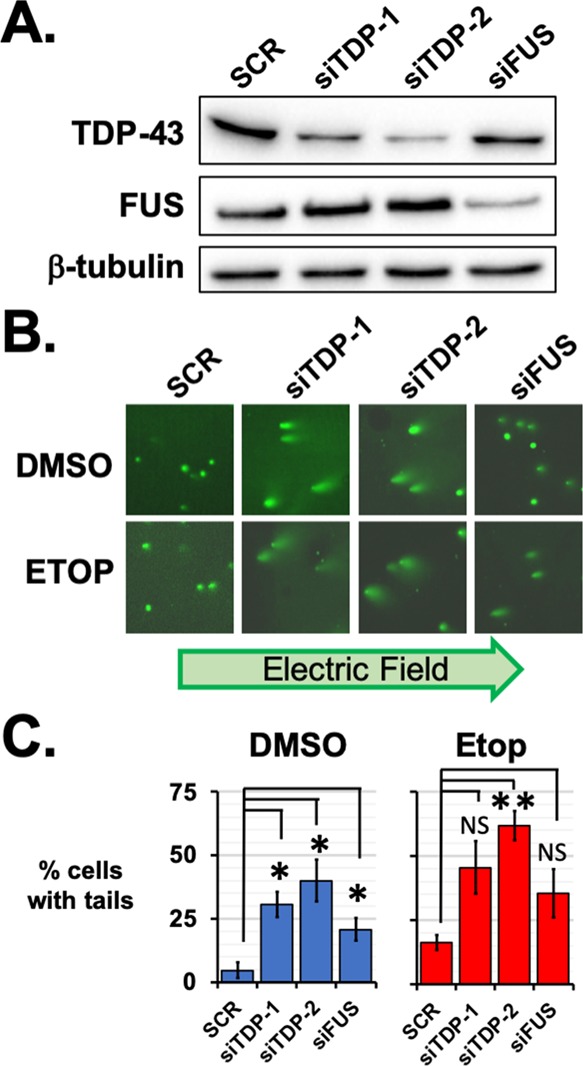
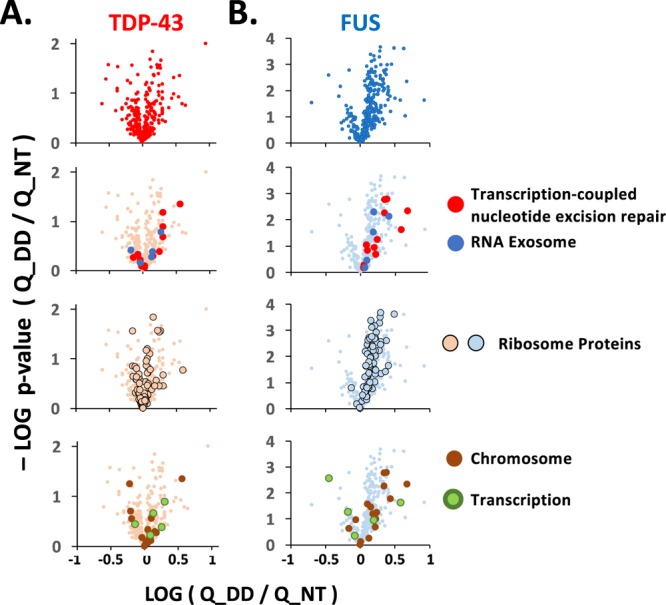
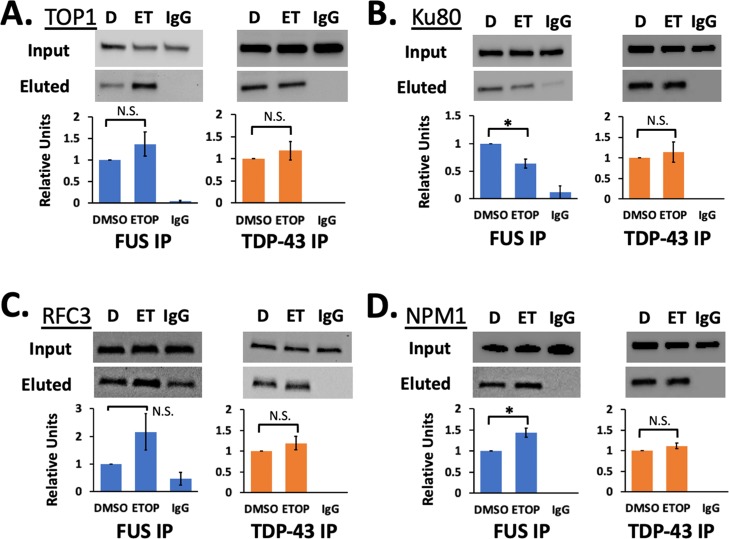
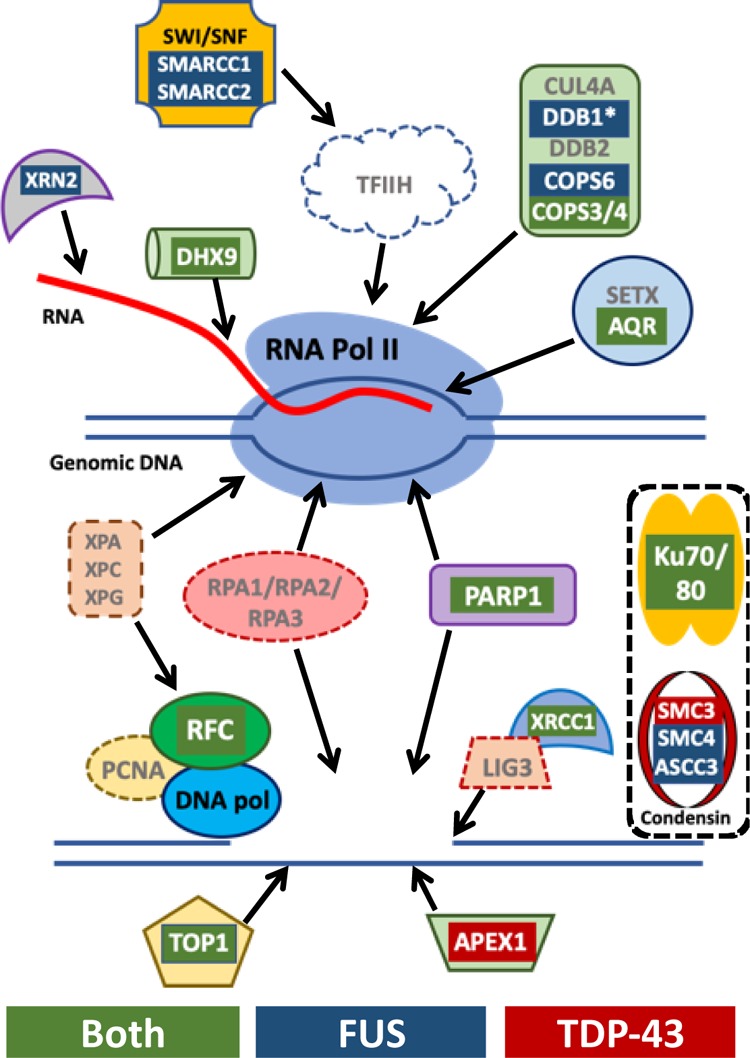
The RNA-binding proteins TDP-43 and FUS are tied as the third leading known genetic cause for amyotrophic lateral sclerosis (ALS), and TDP-43 proteopathies are found in nearly all ALS patients. Both the natural function and contribution to pathology for TDP-43 remain unclear. The intersection of functions between TDP-43 and FUS can focus attention for those natural functions mostly likely to be relevant to disease. Here, we compare the role played by TDP-43 and FUS, maintaining chromatin stability for dividing HEK293T cells. We also determine and compare the interactomes of TDP-43 and FUS, quantitating changes in those before and after DNA damage. Finally, selected interactions with known importance to DNA damage repair were validated by co-immunoprecipitation assays. This study uncovered TDP-43 and FUS binding to several factors important to DNA repair mechanisms that can be replication-dependent, -independent, or both. These results provide further evidence that TDP-43 has an important role in DNA stability and provide new ways that TDP-43 can bind to the machinery that guards DNA integrity in cells.
Keywords: DNA damage repair; FUS; TDP-43; amyotrophic lateral sclerosis; frontal temporal dementia; transcription.
Conflict of interest statement
The authors declare no competing financial interest.
Figures

References
-
- Krach F.; Batra R.; Wheeler E. C.; Vu A. Q.; Wang R.; Hutt K.; Rabin S. J.; Baughn M. W.; Libby R. T.; Diaz-Garcia S.; Stauffer J.; Pirie E.; Saberi S.; Rodriguez M.; Madrigal A. A.; Kohl Z.; Winner B.; Yeo G. W.; Ravits J. Transcriptome-pathology correlation identifies interplay between TDP-43 and the expression of its kinase CK1E in sporadic ALS. Acta Neuropathol. 2018, 136, 405–423. 10.1007/s00401-018-1870-7. - DOI - PMC - PubMed
-
- Kapeli K.; Pratt G. A.; Vu A. Q.; Hutt K. R.; Martinez F. J.; Sundararaman B.; Batra R.; Freese P.; Lambert N. J.; Huelga S. C.; Chun S. J.; Liang T. Y.; Chang J.; Donohue J. P.; Shiue L.; Zhang J.; Zhu H.; Cambi F.; Kasarskis E.; Hoon S.; Ares M. Jr.; Burge C. B.; Ravits J.; Rigo F.; Yeo G. W. Distinct and shared functions of ALS-associated proteins TDP-43, FUS and TAF15 revealed by multisystem analyses. Nat. Commun. 2016, 7, 12143. 10.1038/ncomms12143. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
