

The mammalian spinal commissural system: properties and functions
- PMID: 31693445
- PMCID: PMC6985854
- DOI: 10.1152/jn.00347.2019
The mammalian spinal commissural system: properties and functions
Abstract
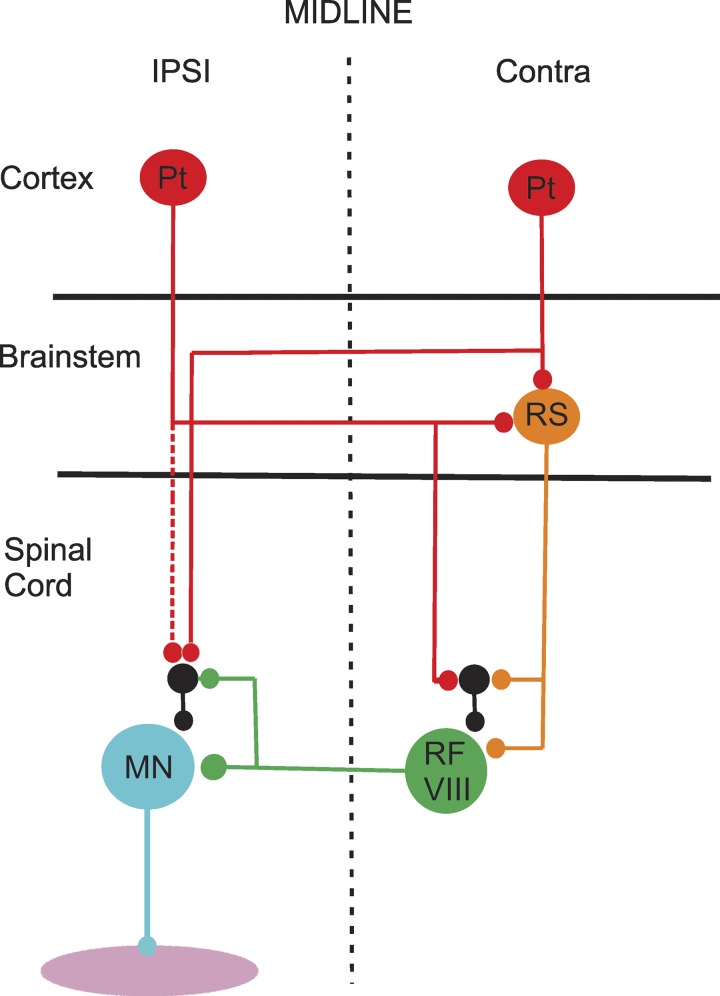
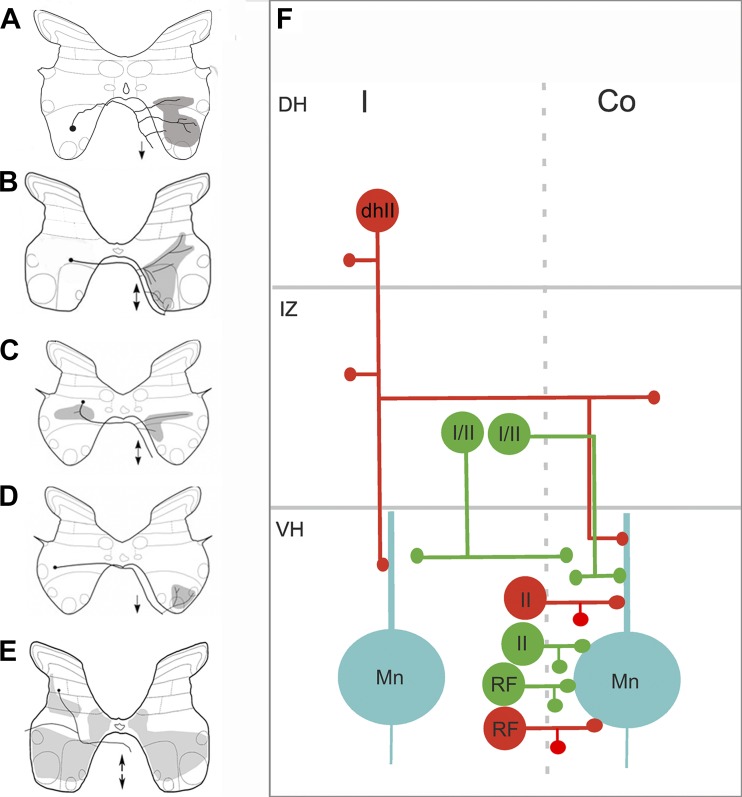
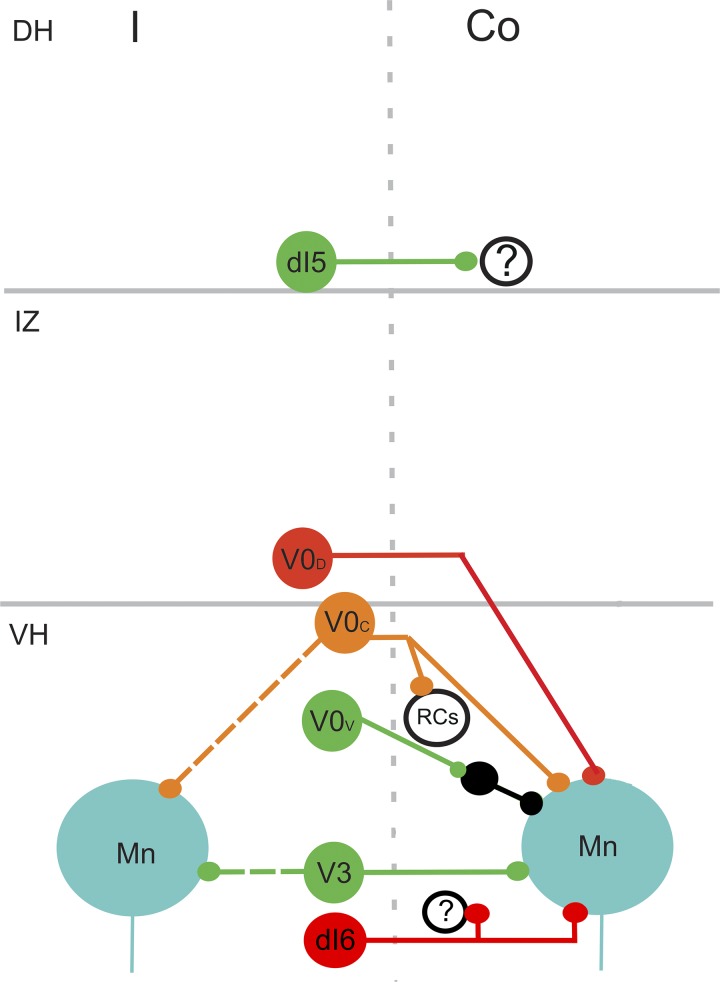
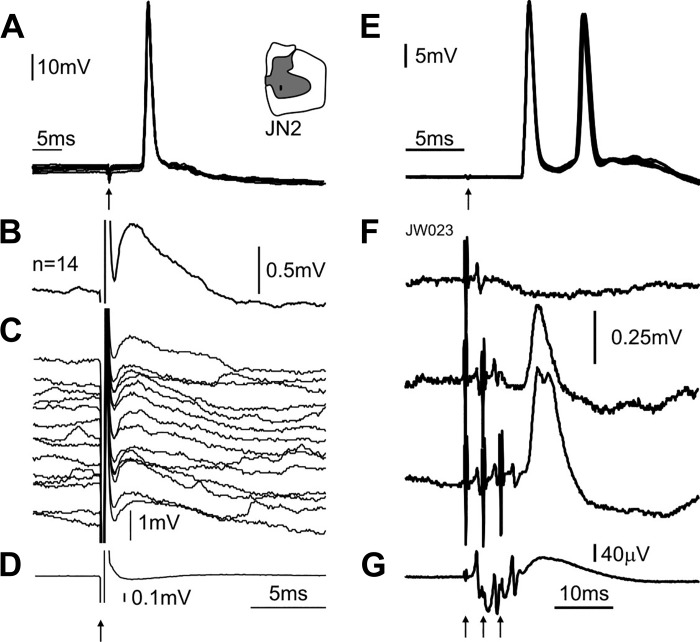
Commissural systems are essential components of motor circuits that coordinate left-right activity of the skeletomuscular system. Commissural systems are found at many levels of the neuraxis including the cortex, brainstem, and spinal cord. In this review we will discuss aspects of the mammalian spinal commissural system. We will focus on commissural interneurons, which project from one side of the cord to the other and form axonal terminations that are confined to the cord itself. Commissural interneurons form heterogeneous populations and influence a variety of spinal circuits. They can be defined according to a variety of criteria including, location in the spinal gray matter, axonal projections and targets, neurotransmitter phenotype, activation properties, and embryological origin. At present, we do not have a comprehensive classification of these cells, but it is clear that cells located within different areas of the gray matter have characteristic properties and make particular contributions to motor circuits. The contribution of commissural interneurons to locomotor function and posture is well established and briefly discussed. However, their role in other goal-orientated behaviors such as grasping, reaching, and bimanual tasks is less clear. This is partly because we only have limited information about the organization and functional properties of commissural interneurons in the cervical spinal cord of primates, including humans. In this review we shall discuss these various issues. First, we will consider the properties of commissural interneurons and subsequently examine what is known about their functions. We then discuss how they may contribute to restoration of function following spinal injury and stroke.
Keywords: bilateral; commissural; movement.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
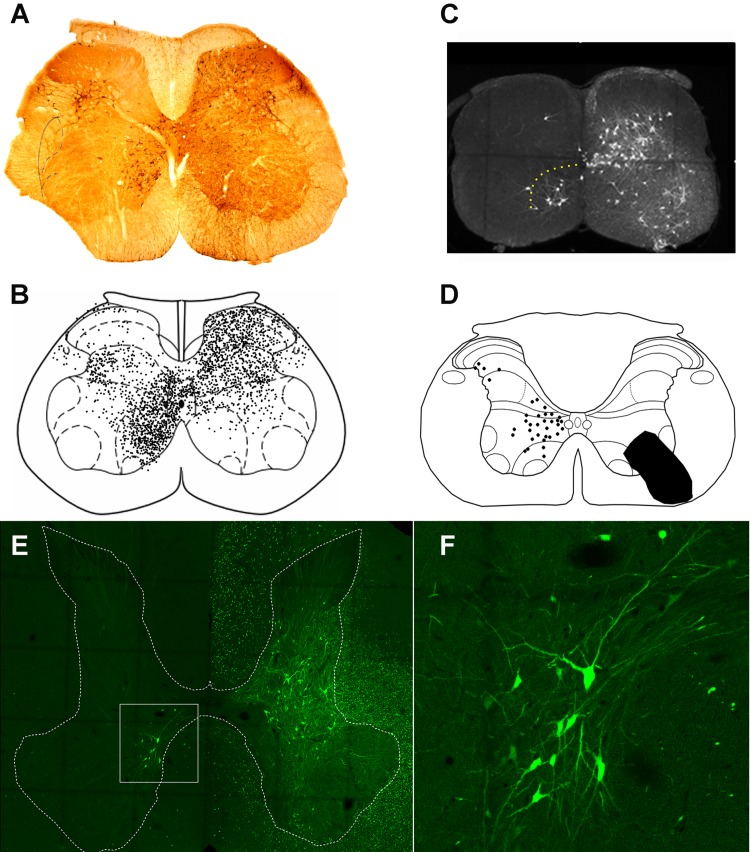
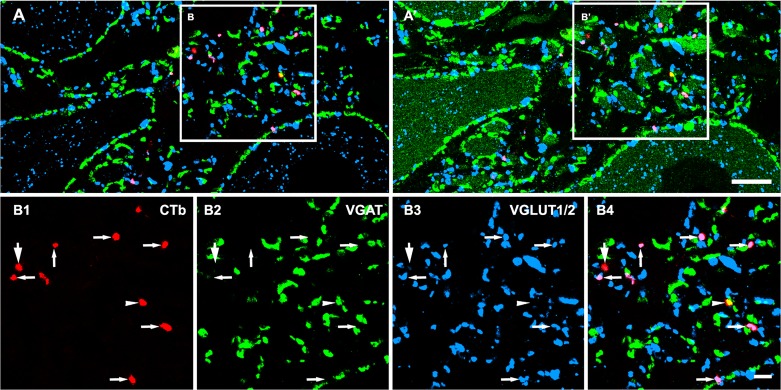
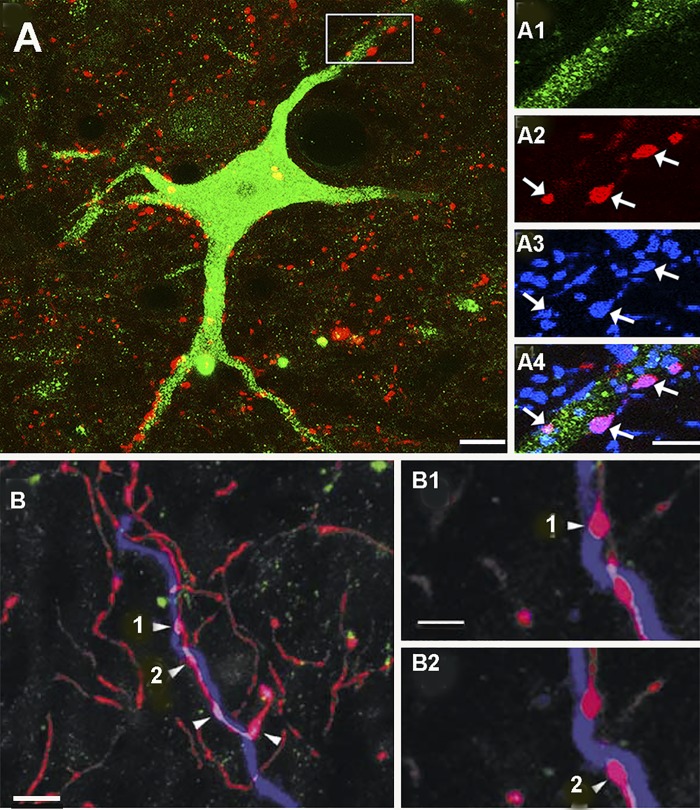
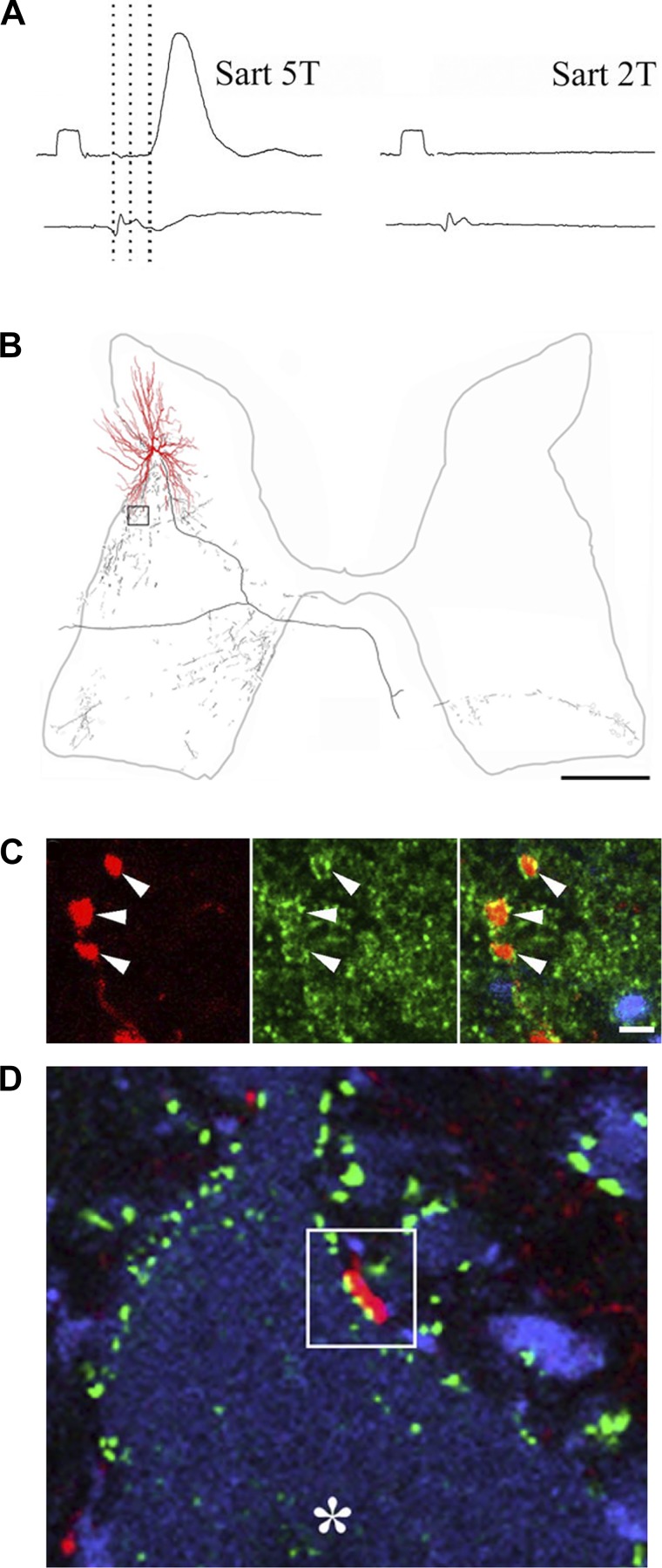
References
-
- Alstermark B, Kümmel H. Transneuronal transport of wheat germ agglutinin conjugated horseradish peroxidase into last order spinal interneurones projecting to acromio- and spinodeltoideus motoneurones in the cat. 1. Location of labelled interneurones and influence of synaptic activity on the transneuronal transport. Exp Brain Res 80: 83–95, 1990. doi: 10.1007/BF00228850. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical