

Metformin Improves Mitochondrial Respiratory Activity through Activation of AMPK
- PMID: 31693892
- PMCID: PMC6866677
- DOI: 10.1016/j.celrep.2019.09.070
Metformin Improves Mitochondrial Respiratory Activity through Activation of AMPK
Abstract
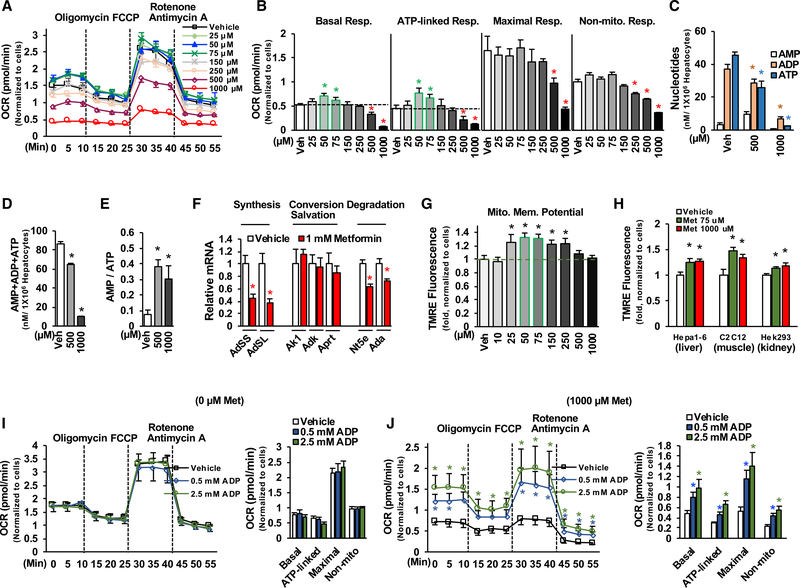
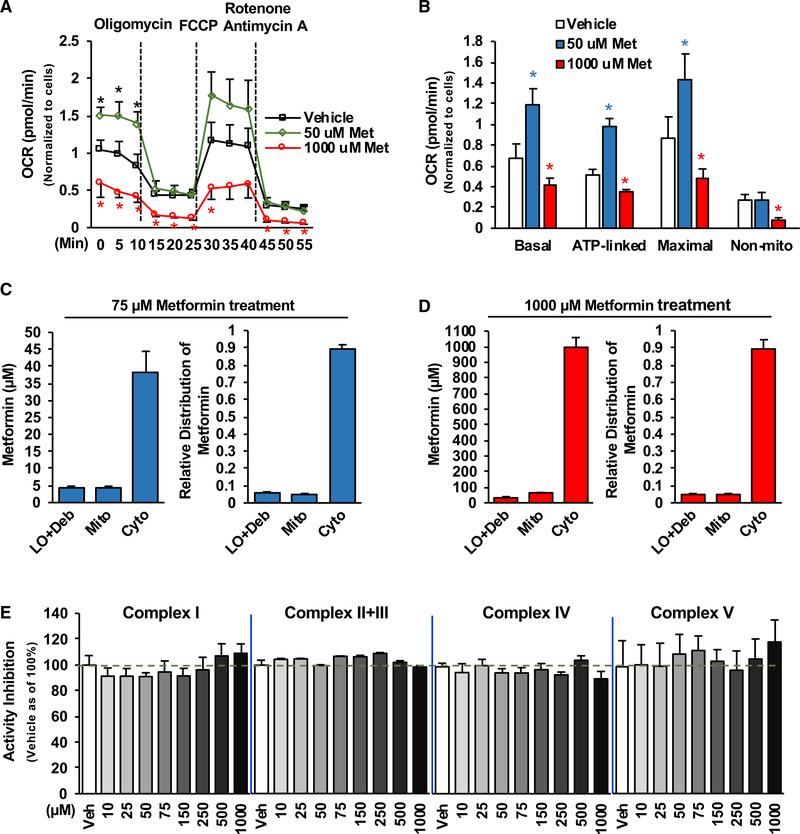
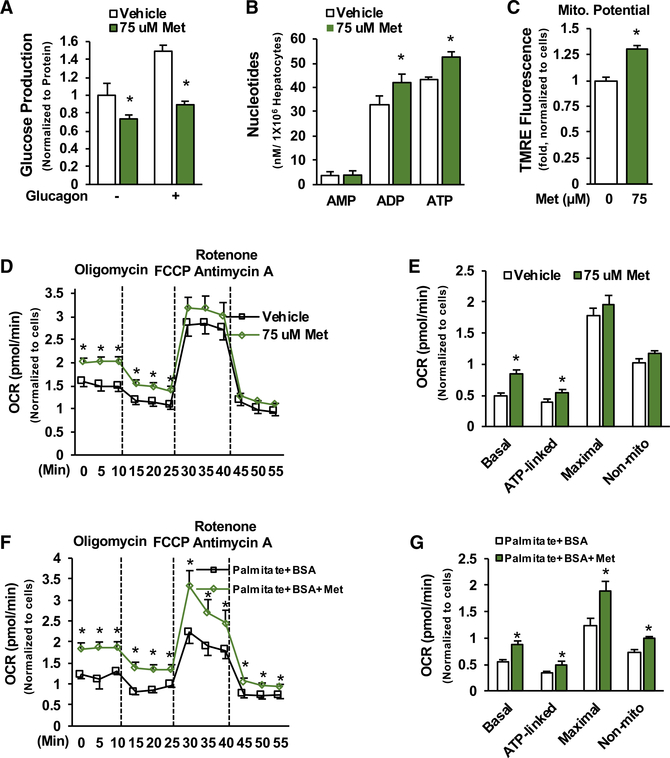
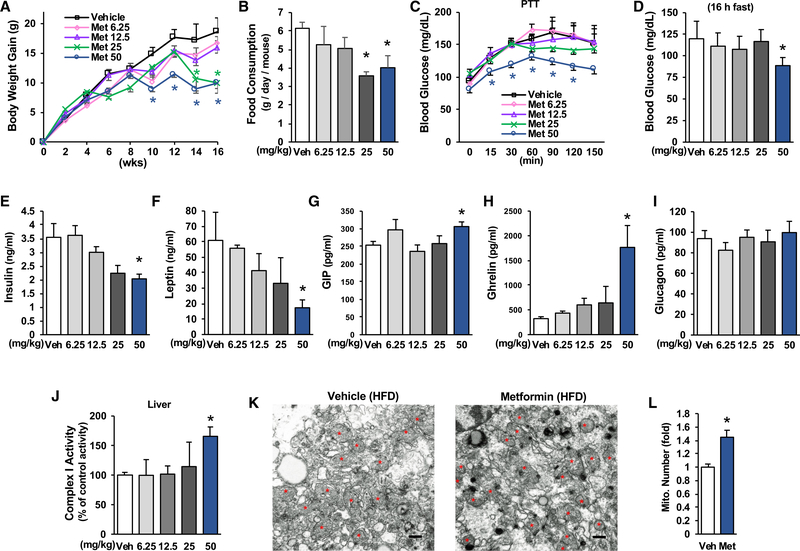
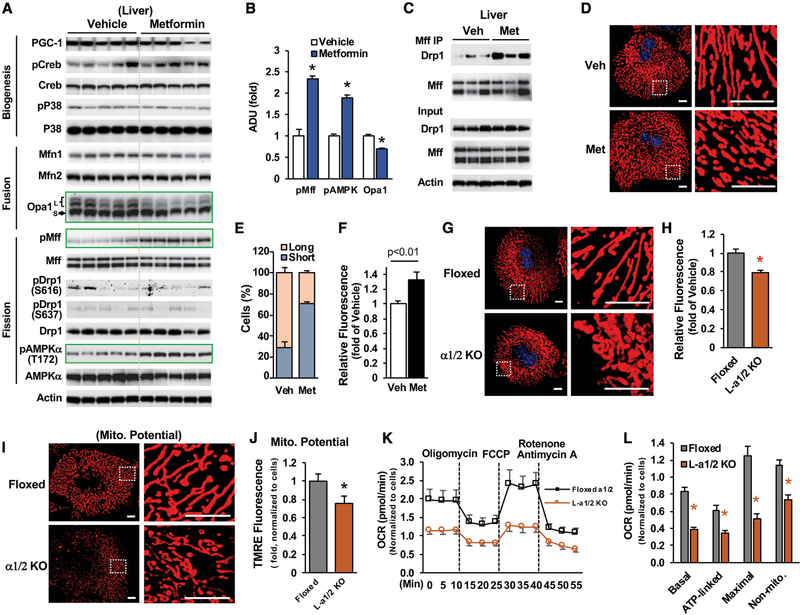
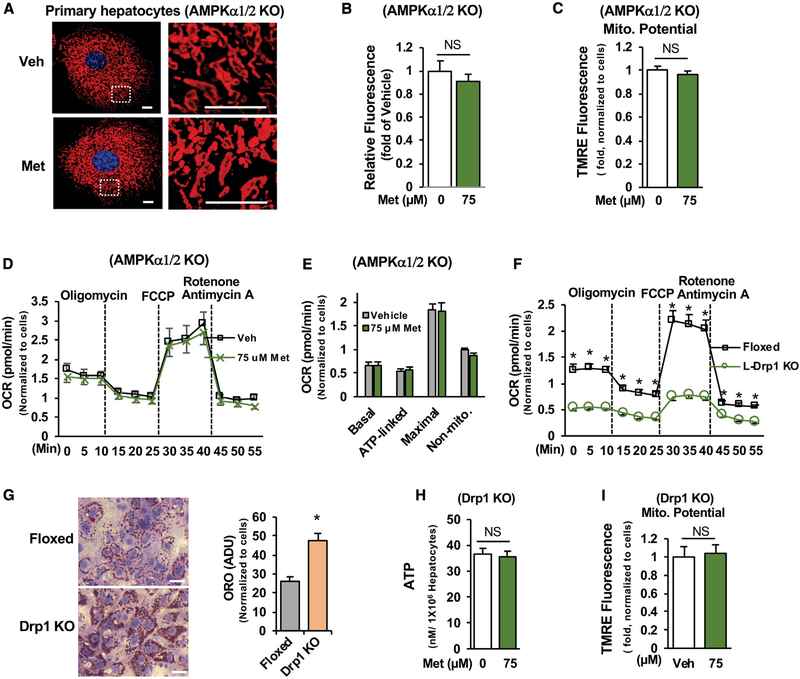
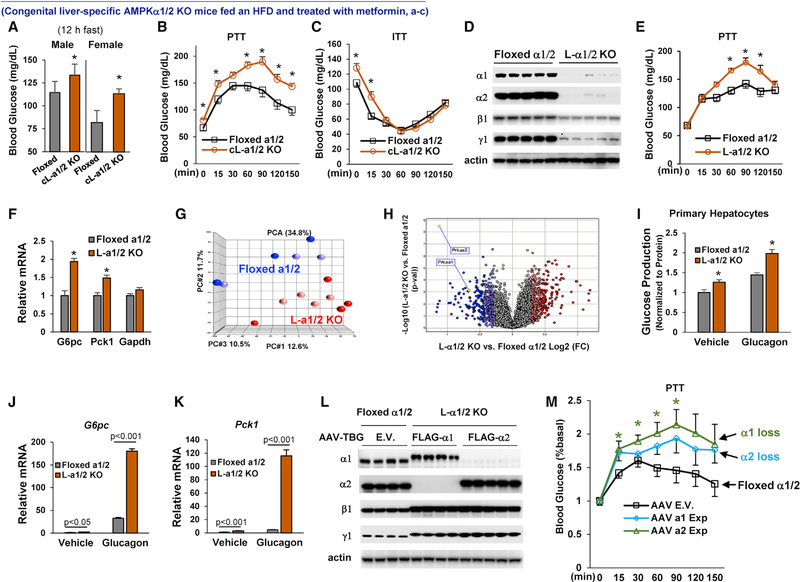
Impaired mitochondrial respiratory activity contributes to the development of insulin resistance in type 2 diabetes. Metformin, a first-line antidiabetic drug, functions mainly by improving patients' hyperglycemia and insulin resistance. However, its mechanism of action is still not well understood. We show here that pharmacological metformin concentration increases mitochondrial respiration, membrane potential, and ATP levels in hepatocytes and a clinically relevant metformin dose increases liver mitochondrial density and complex 1 activity along with improved hyperglycemia in high-fat- diet (HFD)-fed mice. Metformin, functioning through 5' AMP-activated protein kinase (AMPK), promotes mitochondrial fission to improve mitochondrial respiration and restore the mitochondrial life cycle. Furthermore, HFD-fed-mice with liver-specific knockout of AMPKα1/2 subunits exhibit higher blood glucose levels when treated with metformin. Our results demonstrate that activation of AMPK by metformin improves mitochondrial respiration and hyperglycemia in obesity. We also found that supra-pharmacological metformin concentrations reduce adenine nucleotides, resulting in the halt of mitochondrial respiration. These findings suggest a mechanism for metformin's anti-tumor effects.
Keywords: AMPK; Drp1; adenine nucleotides; diabetes; insulin resistance; membrane potential; metformin; mitochondrial respiration/fission.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests
Figures
Comment in
-
Mitochondrial Complex I Inhibition by Metformin: Drug-Exercise Interactions.Trends Endocrinol Metab. 2020 Apr;31(4):269-271. doi: 10.1016/j.tem.2020.02.003. Epub 2020 Feb 20. Trends Endocrinol Metab. 2020. PMID: 32187522
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
