

AMIGO2 Scales Dendrite Arbors in the Retina
- PMID: 31693896
- PMCID: PMC6871773
- DOI: 10.1016/j.celrep.2019.09.085
AMIGO2 Scales Dendrite Arbors in the Retina
Abstract
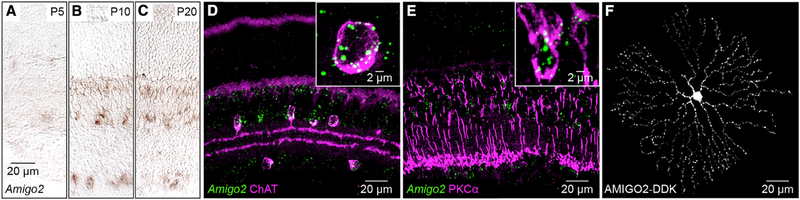
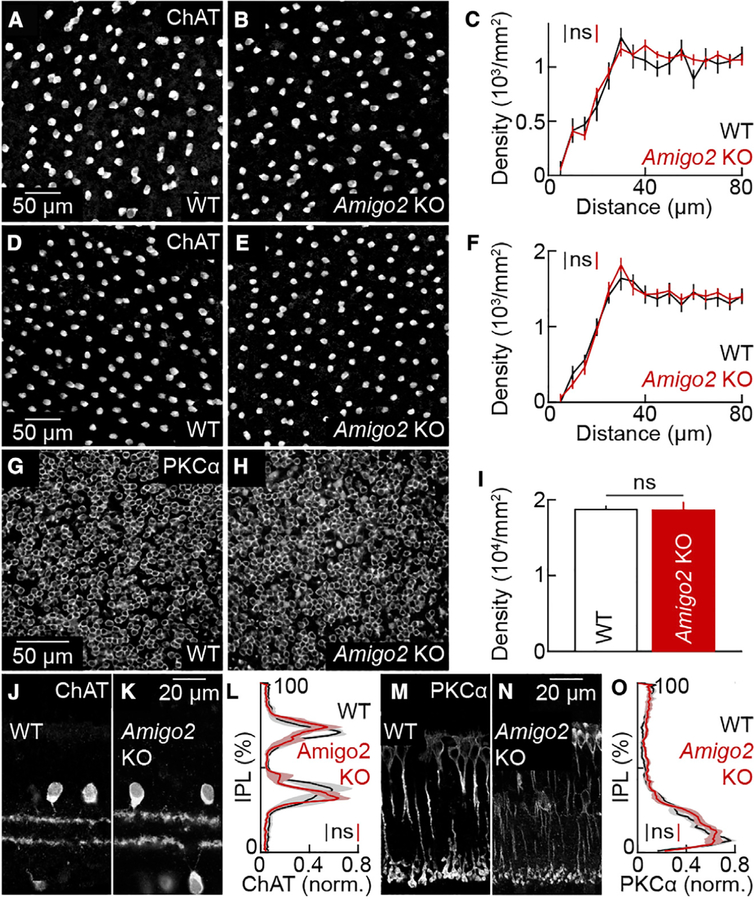
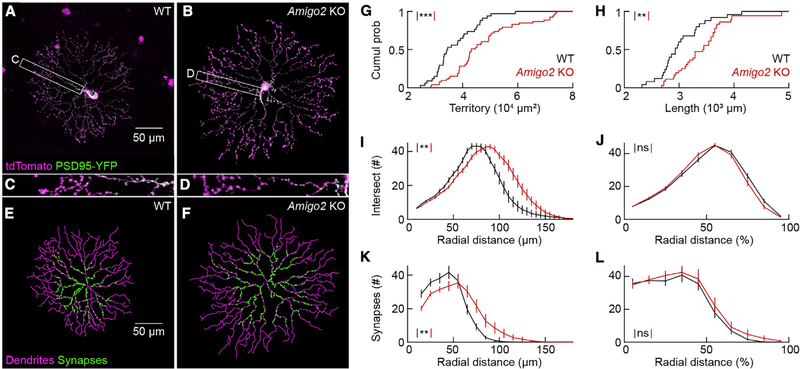
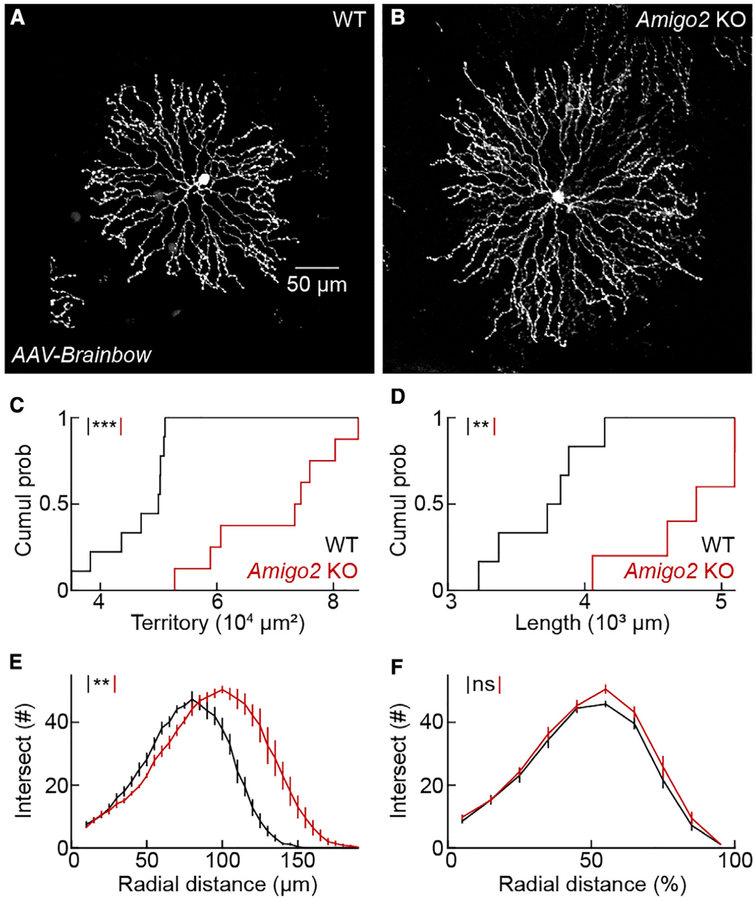
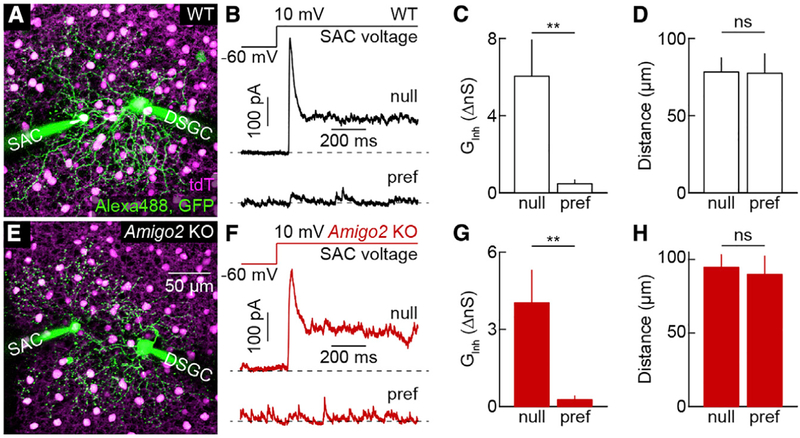
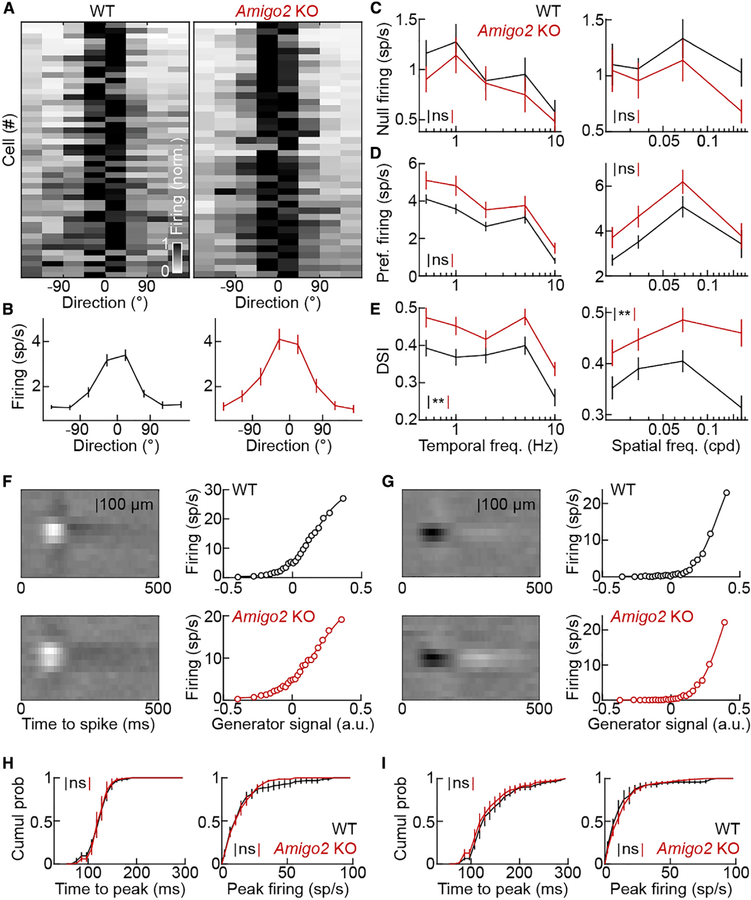
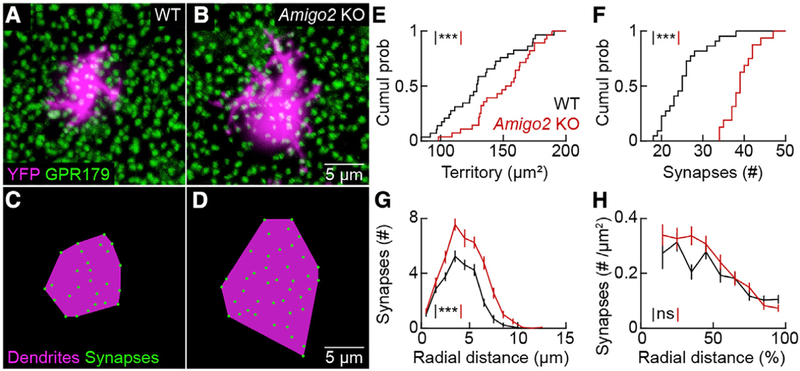
The size of dendrite arbors shapes their function and differs vastly between neuron types. The signals that control dendritic arbor size remain obscure. Here, we find that in the retina, starburst amacrine cells (SACs) and rod bipolar cells (RBCs) express the homophilic cell-surface protein AMIGO2. In Amigo2 knockout (KO) mice, SAC and RBC dendrites expand while arbors of other retinal neurons remain stable. SAC dendrites are divided into a central input region and a peripheral output region that provides asymmetric inhibition to direction-selective ganglion cells (DSGCs). Input and output compartments scale precisely with increased arbor size in Amigo2 KO mice, and SAC dendrites maintain asymmetric connectivity with DSGCs. Increased coverage of SAC dendrites is accompanied by increased direction selectivity of DSGCs without changes to other ganglion cells. Our results identify AMIGO2 as a cell-type-specific dendritic scaling factor and link dendrite size and coverage to visual feature detection.
Keywords: LRR; dendrites; development; direction selectivity; leucine-rich repeat protein; retina; rod bipolar cell; starburst amacrine cell.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Briggman KL, Helmstaedter M, and Denk W (2011). Wiring specificity in the direction-selectivity circuit of the retina. Nature 471, 183–188. - PubMed
-
- Brown SP, He S, and Masland RH (2000). Receptive field microstructure and dendritic geometry of retinal ganglion cells. Neuron 27, 371–383. - PubMed
-
- Cai D, Cohen KB, Luo T, Lichtman JW, and Sanes JR (2013). Improved tools for the Brainbow toolbox. Nat. Methods 10, 540–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
