

Adipose tissue NAD+ biosynthesis is required for regulating adaptive thermogenesis and whole-body energy homeostasis in mice
- PMID: 31694884
- PMCID: PMC6876243
- DOI: 10.1073/pnas.1909917116
Adipose tissue NAD+ biosynthesis is required for regulating adaptive thermogenesis and whole-body energy homeostasis in mice
Abstract
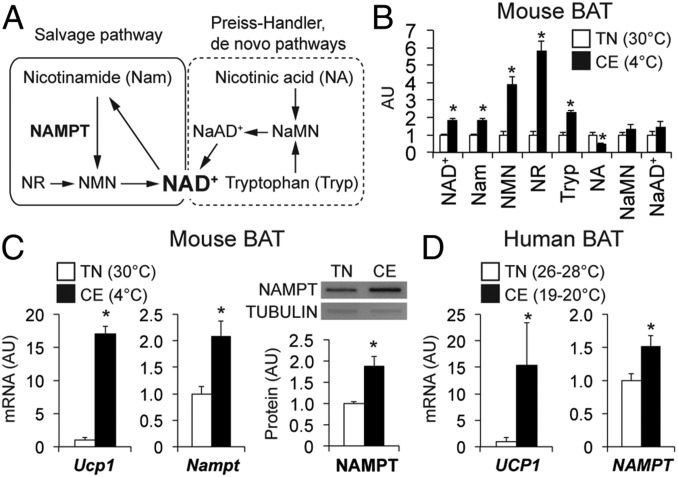
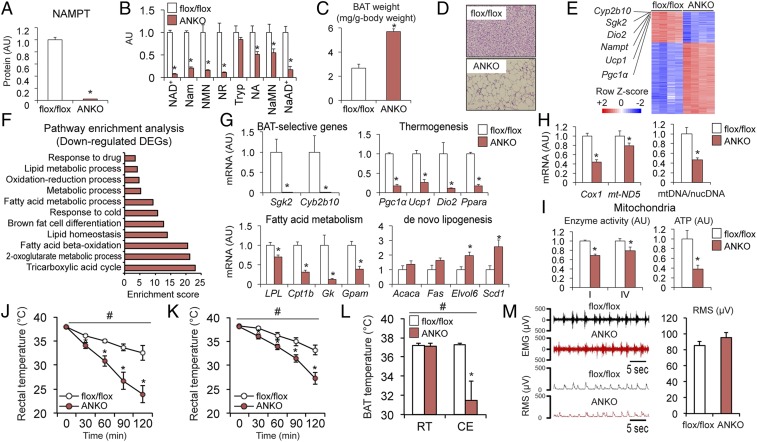
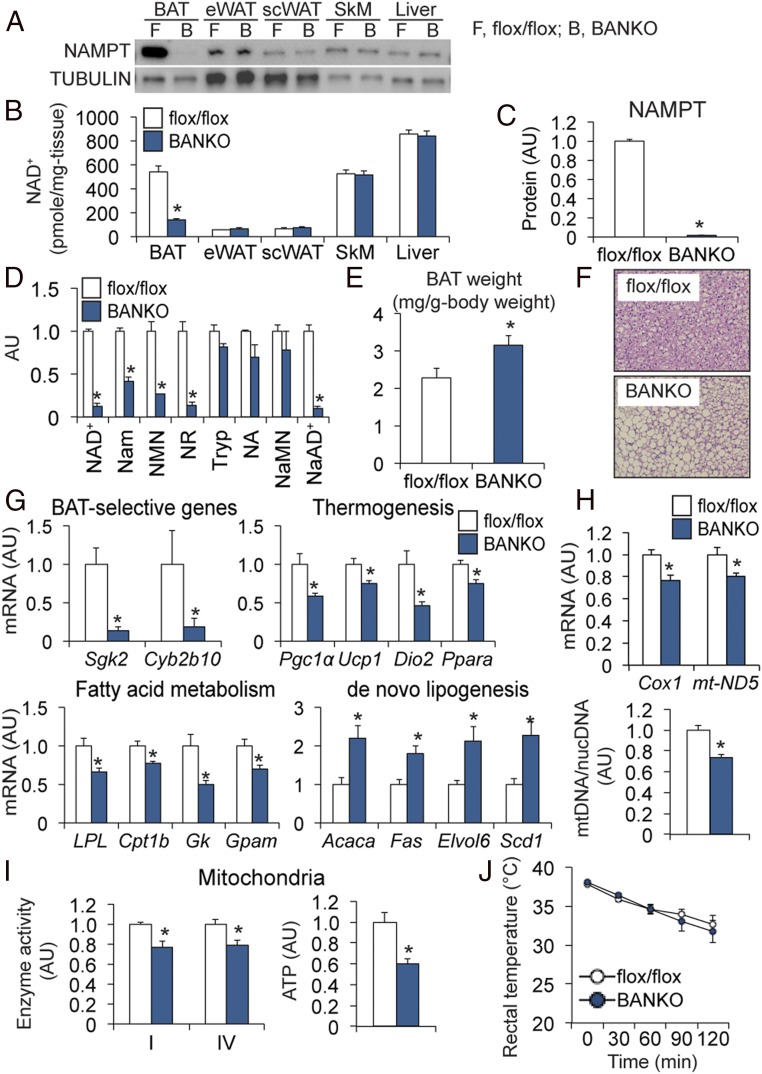
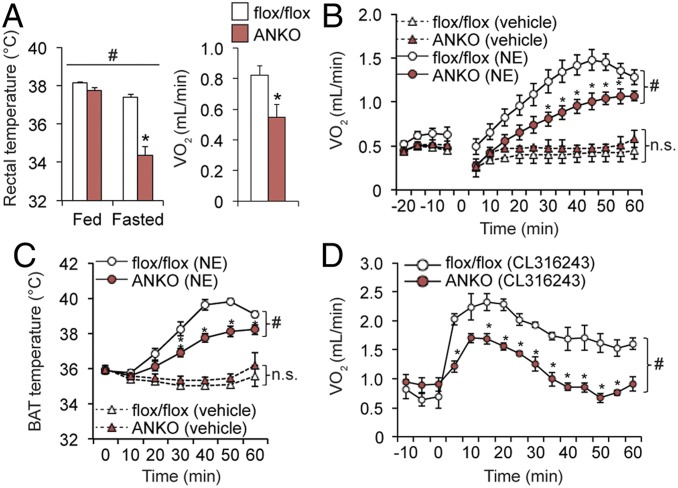
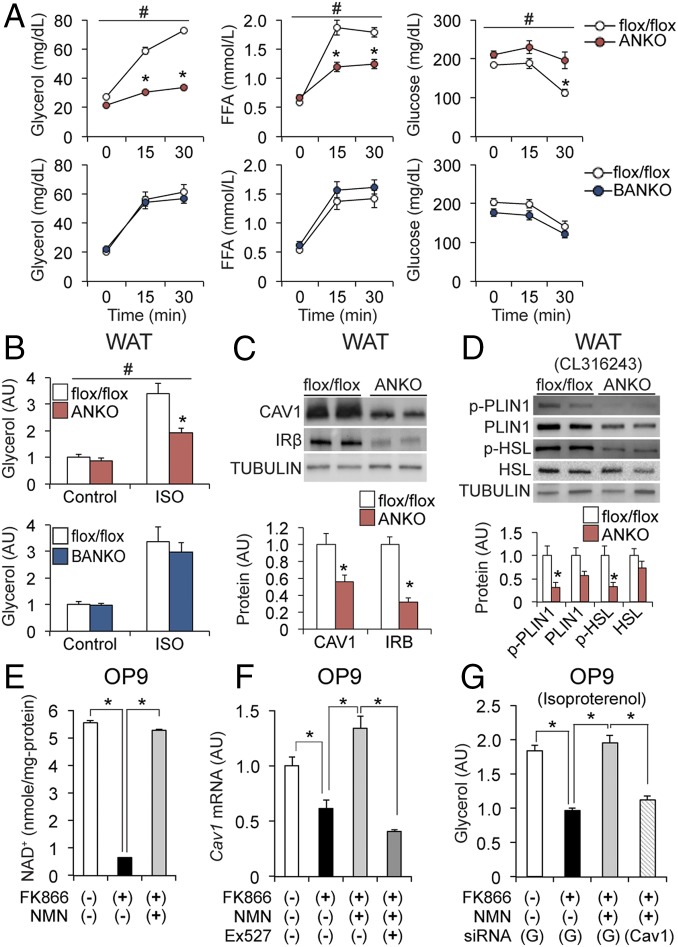
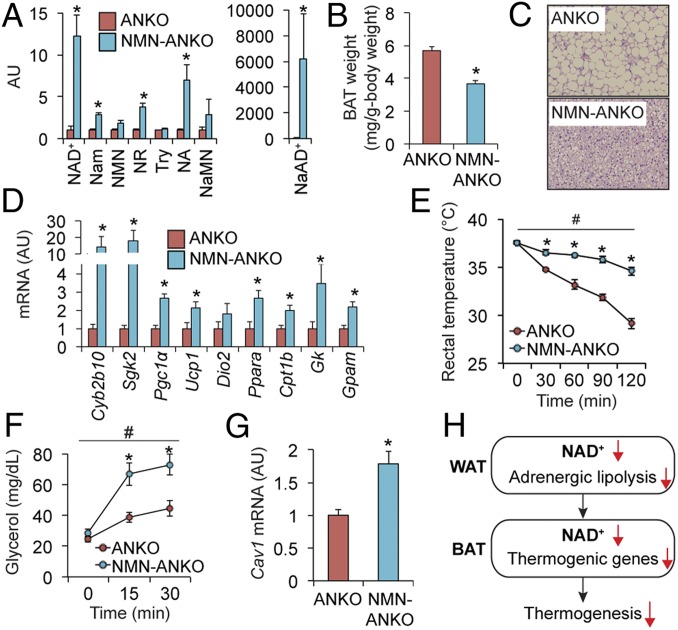
Nicotinamide adenine dinucleotide (NAD+) is a critical coenzyme for cellular energy metabolism. The aim of the present study was to determine the importance of brown and white adipose tissue (BAT and WAT) NAD+ metabolism in regulating whole-body thermogenesis and energy metabolism. Accordingly, we generated and analyzed adipocyte-specific nicotinamide phosphoribosyltransferase (Nampt) knockout (ANKO) and brown adipocyte-specific Nampt knockout (BANKO) mice because NAMPT is the rate-limiting NAD+ biosynthetic enzyme. We found ANKO mice, which lack NAMPT in both BAT and WAT, had impaired gene programs involved in thermogenesis and mitochondrial function in BAT and a blunted thermogenic (rectal temperature, BAT temperature, and whole-body oxygen consumption) response to acute cold exposure, prolonged fasting, and administration of β-adrenergic agonists (norepinephrine and CL-316243). In addition, the absence of NAMPT in WAT markedly reduced adrenergic-mediated lipolytic activity, likely through inactivation of the NAD+-SIRT1-caveolin-1 axis, which limits an important fuel source fatty acid for BAT thermogenesis. These metabolic abnormalities were rescued by treatment with nicotinamide mononucleotide (NMN), which bypasses the block in NAD+ synthesis induced by NAMPT deficiency. Although BANKO mice, which lack NAMPT in BAT only, had BAT cellular alterations similar to the ANKO mice, BANKO mice had normal thermogenic and lipolytic responses. We also found NAMPT expression in supraclavicular adipose tissue (where human BAT is localized) obtained from human subjects increased during cold exposure, suggesting our finding in rodents could apply to people. These results demonstrate that adipose NAMPT-mediated NAD+ biosynthesis is essential for regulating adaptive thermogenesis, lipolysis, and whole-body energy metabolism.
Keywords: NAD; adipose tissue; energy metabolism; lipolysis; thermogenesis.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
