

Murine Coronavirus Infection Activates the Aryl Hydrocarbon Receptor in an Indoleamine 2,3-Dioxygenase-Independent Manner, Contributing to Cytokine Modulation and Proviral TCDD-Inducible-PARP Expression
- PMID: 31694960
- PMCID: PMC7000979
- DOI: 10.1128/JVI.01743-19
Murine Coronavirus Infection Activates the Aryl Hydrocarbon Receptor in an Indoleamine 2,3-Dioxygenase-Independent Manner, Contributing to Cytokine Modulation and Proviral TCDD-Inducible-PARP Expression
Abstract
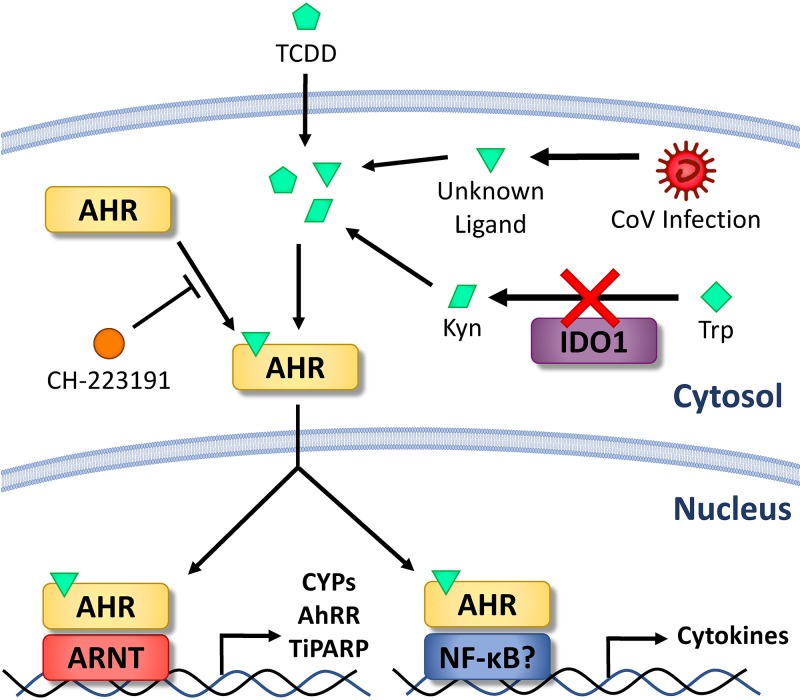
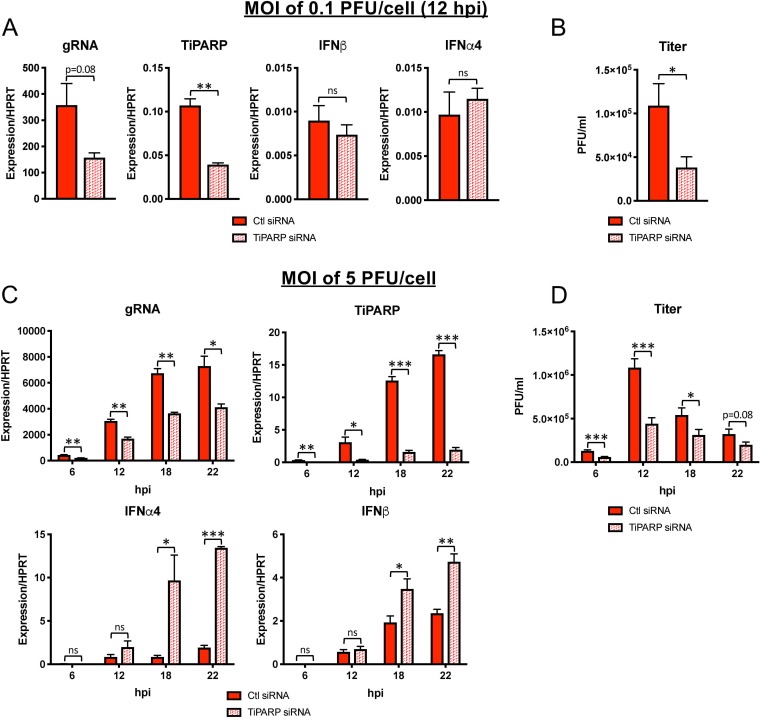
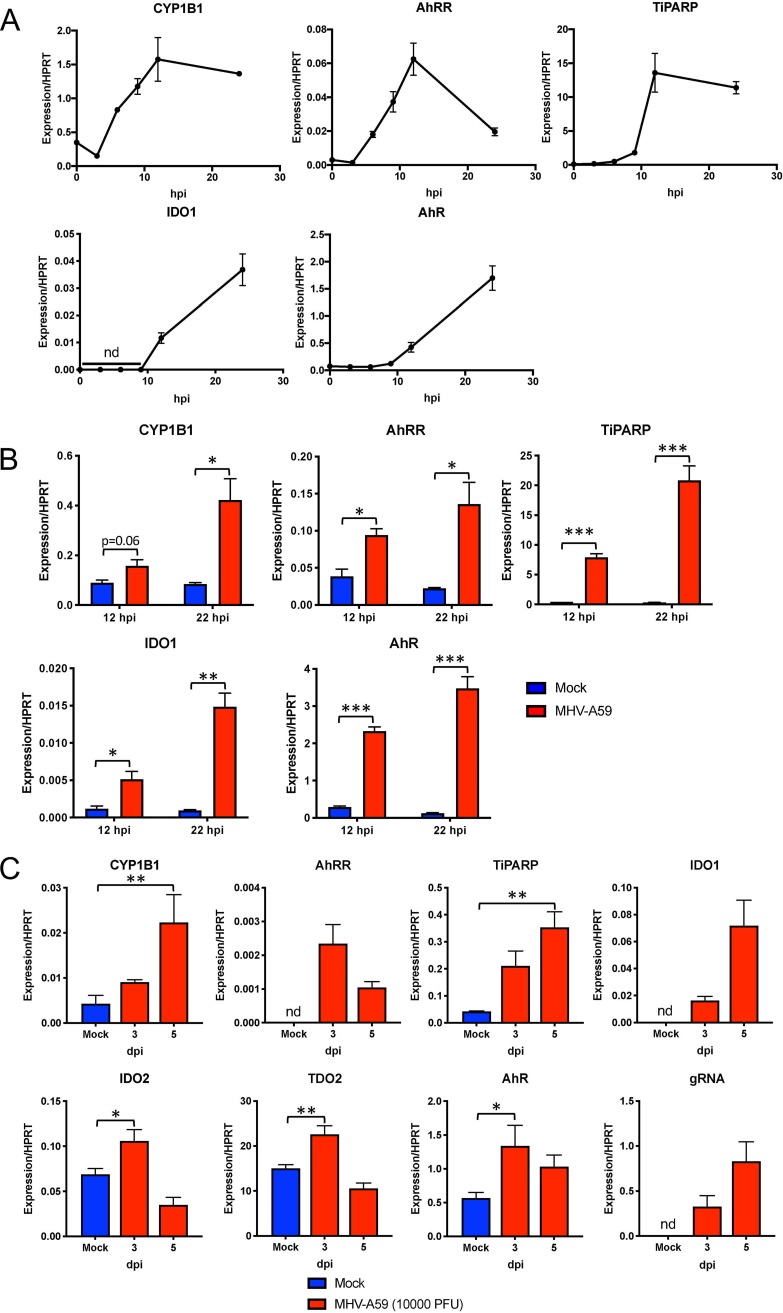
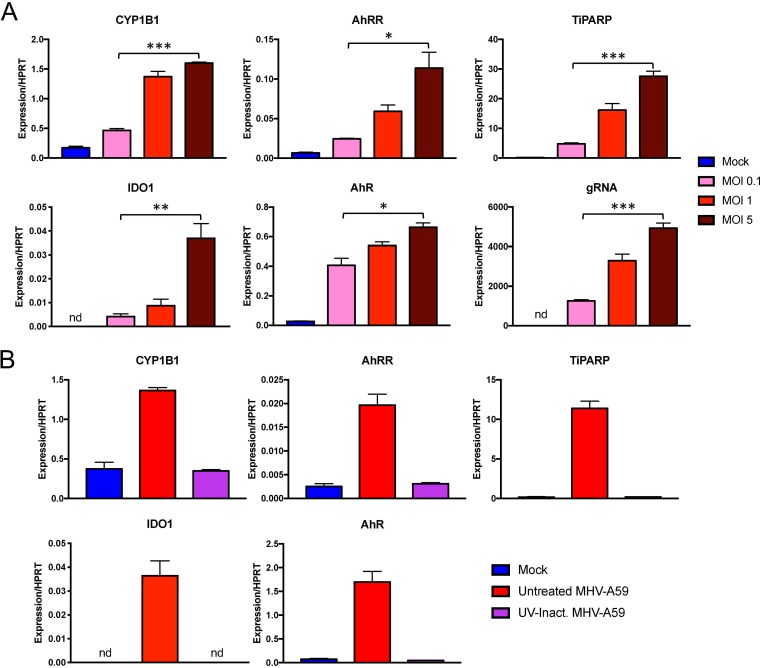
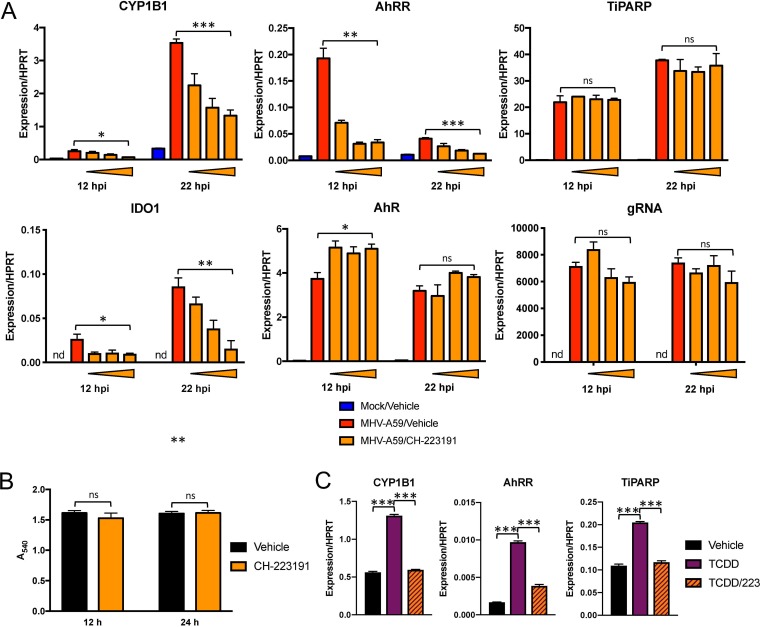
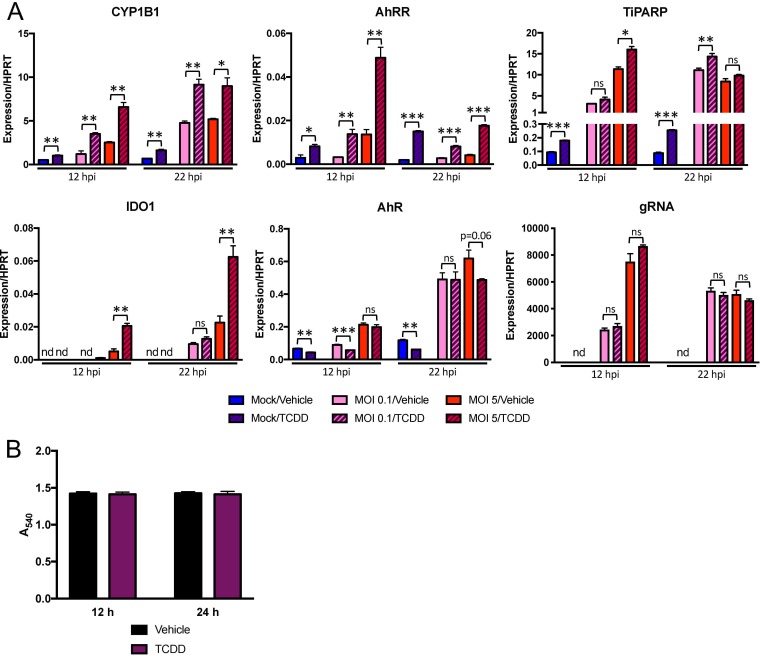
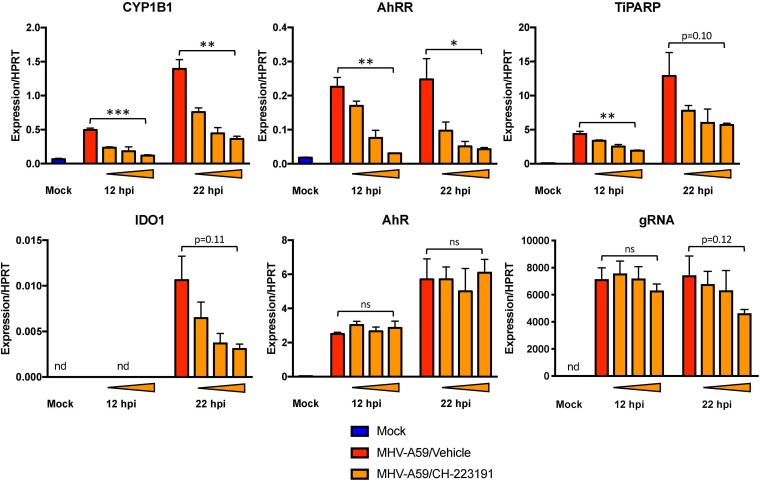
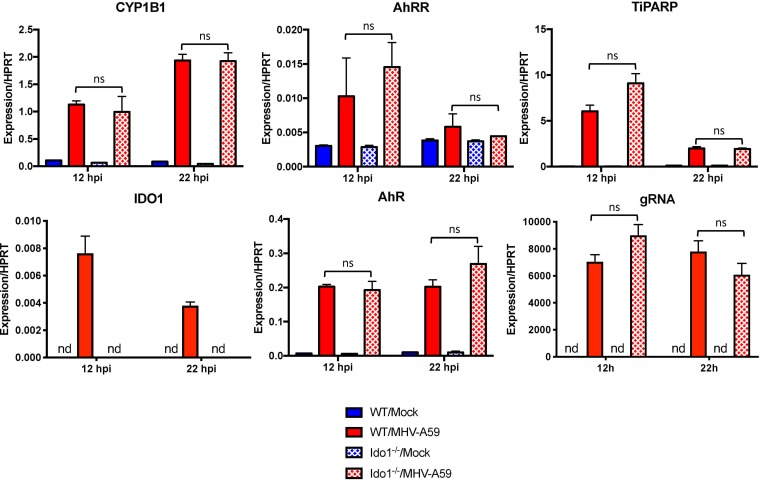
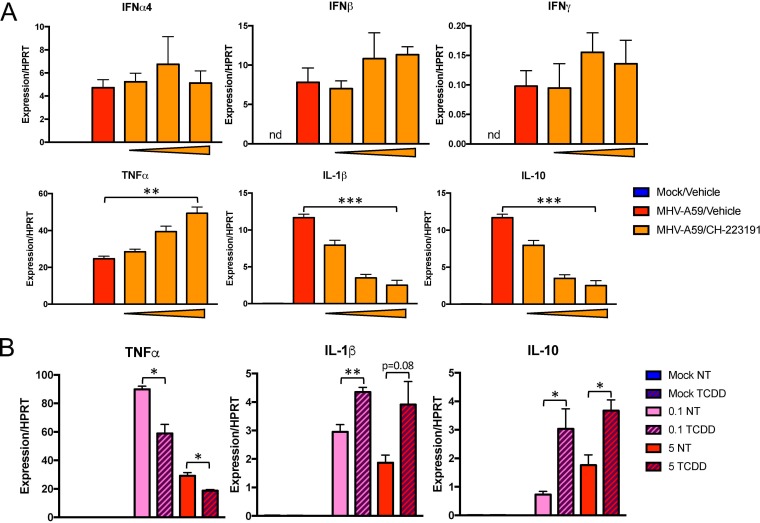
The aryl hydrocarbon receptor (AhR) is a cytoplasmic receptor/transcription factor that modulates several cellular and immunological processes following activation by pathogen-associated stimuli, though its role during virus infection is largely unknown. Here, we show that AhR is activated in cells infected with mouse hepatitis virus (MHV), a coronavirus (CoV), and contributes to the upregulation of downstream effector TCDD-inducible poly(ADP-ribose) polymerase (TiPARP) during infection. Knockdown of TiPARP reduced viral replication and increased interferon expression, suggesting that TiPARP functions in a proviral manner during MHV infection. We also show that MHV replication induced the expression of other genes known to be downstream of AhR in macrophages and dendritic cells and in livers of infected mice. Further, we found that chemically inhibiting or activating AhR reciprocally modulated the expression levels of cytokines induced by infection, specifically, interleukin 1β (IL-1β), IL-10, and tumor necrosis factor alpha (TNF-α), consistent with a role for AhR activation in the host response to MHV infection. Furthermore, while indoleamine 2,3-dioxygenase (IDO1) drives AhR activation in other settings, MHV infection induced equal expression of downstream genes in wild-type (WT) and IDO1-/- macrophages, suggesting an alternative pathway of AhR activation. In summary, we show that coronaviruses elicit AhR activation by an IDO1-independent pathway, contributing to upregulation of downstream effectors, including the proviral factor TiPARP, and to modulation of cytokine gene expression, and we identify a previously unappreciated role for AhR signaling in CoV pathogenesis.IMPORTANCE Coronaviruses are a family of positive-sense RNA viruses with human and agricultural significance. Characterizing the mechanisms by which coronavirus infection dictates pathogenesis or counters the host immune response would provide targets for the development of therapeutics. Here, we show that the aryl hydrocarbon receptor (AhR) is activated in cells infected with a prototypic coronavirus, mouse hepatitis virus (MHV), resulting in the expression of several effector genes. AhR is important for modulation of the host immune response to MHV and plays a role in the expression of TiPARP, which we show is required for maximal viral replication. Taken together, our findings highlight a previously unidentified role for AhR in regulating coronavirus replication and the immune response to the virus.
Keywords: AhR; IDO1; TiPARP; coronavirus; mouse hepatitis virus.
Copyright © 2020 American Society for Microbiology.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
