

Long noncoding RNA DNM3OS promotes prostate stromal cells transformation via the miR-29a/29b/COL3A1 and miR-361/TGFβ1 axes
- PMID: 31694982
- PMCID: PMC6874426
- DOI: 10.18632/aging.102395
Long noncoding RNA DNM3OS promotes prostate stromal cells transformation via the miR-29a/29b/COL3A1 and miR-361/TGFβ1 axes
Abstract
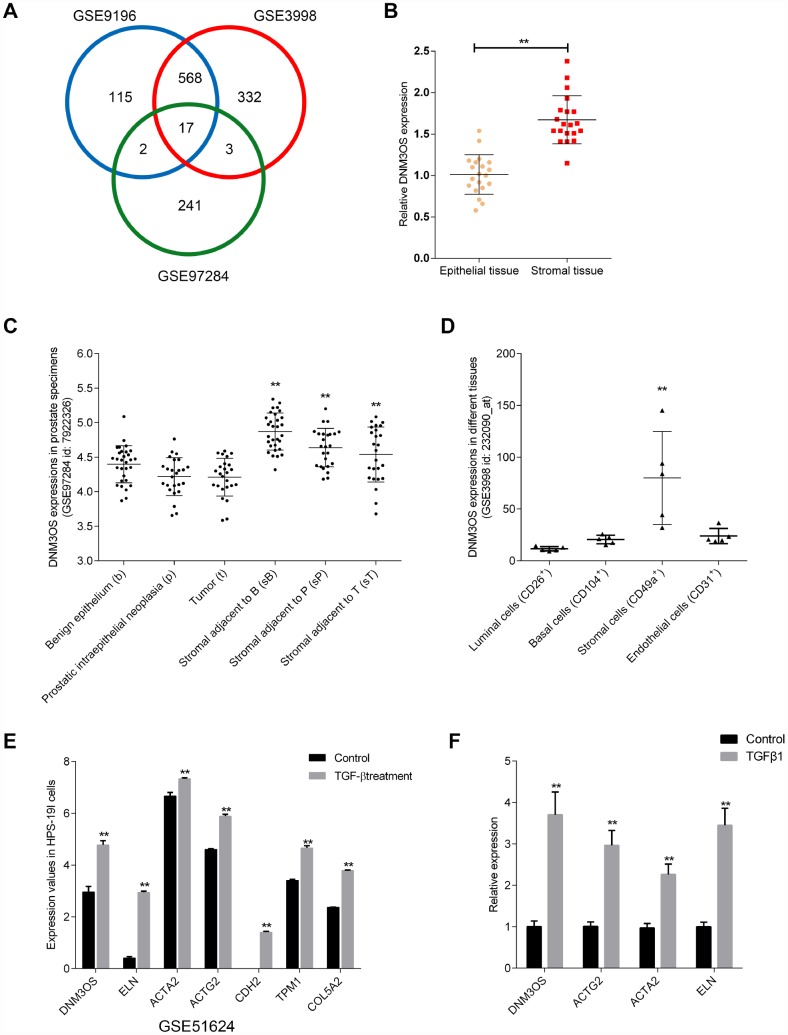
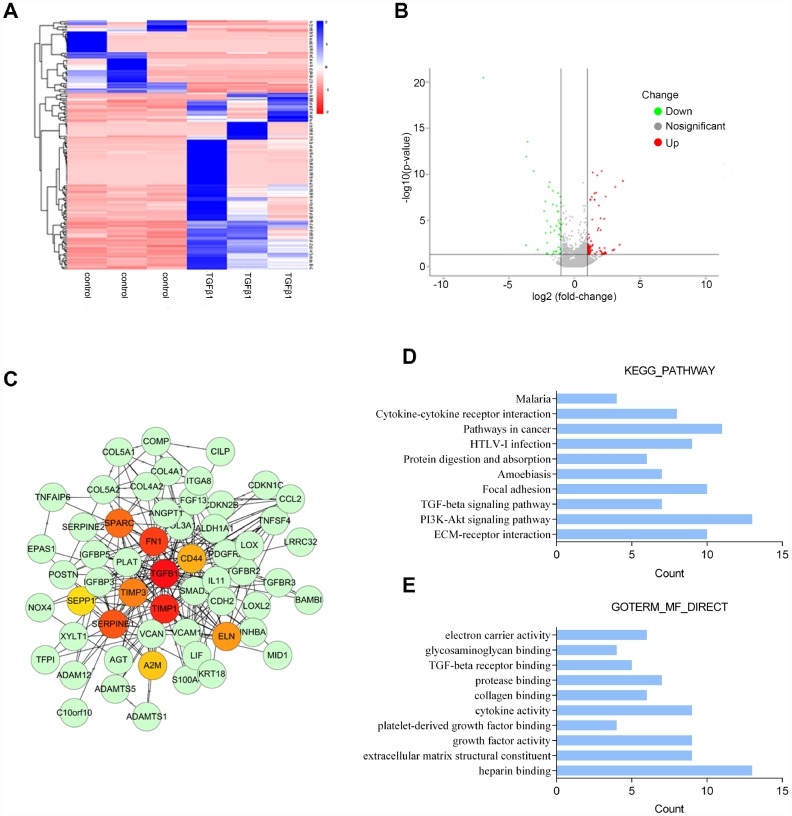
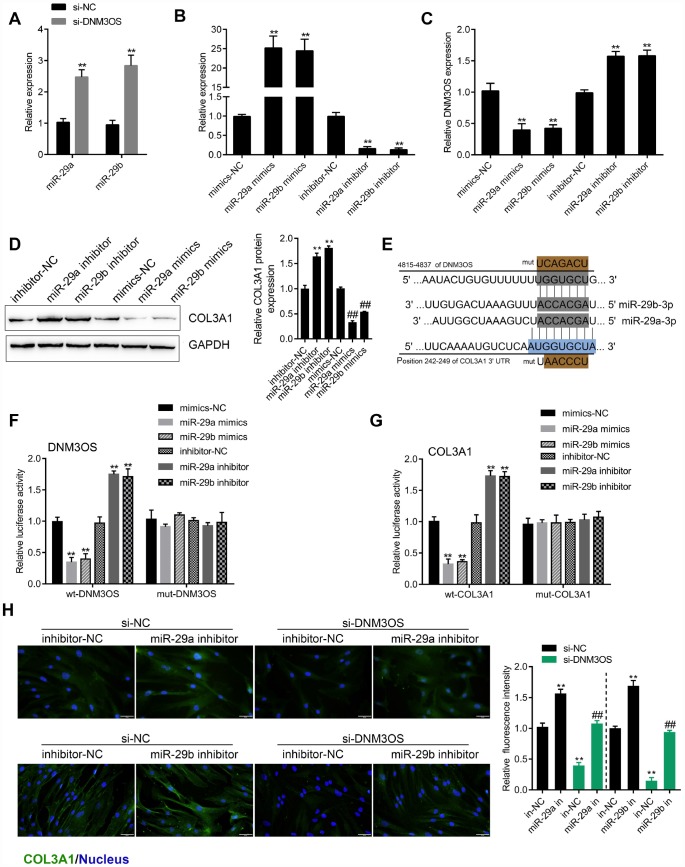
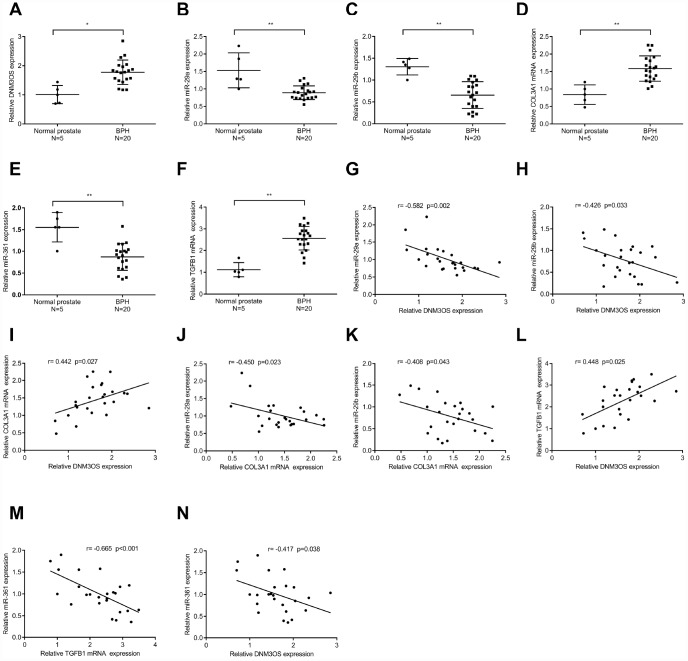
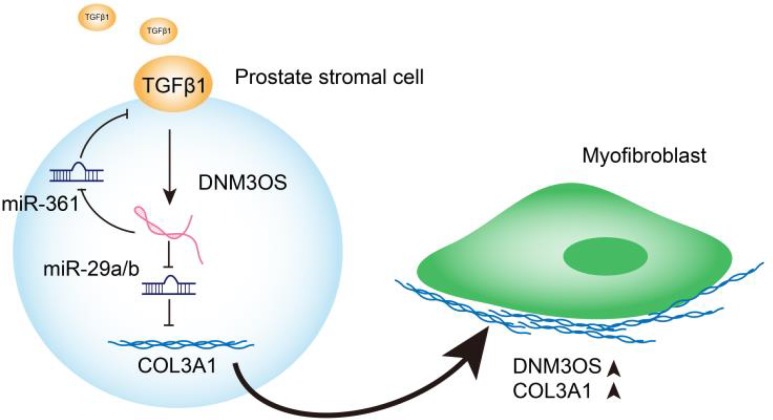
Transforming growth factor-β1 (TGFβ1)-induced differentiation into and the activation of myofibroblasts have been regarded as critical events in benign prostatic hyperplasia (BPH); however, the underlying mechanisms of BPH pathogenesis remain unclear. Microarray profiling, STRING analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway annotation, and Gene Ontology (GO) enrichment analysis were performed to confirm the candidate genes and long non-coding RNA (lncRNAs) related to BPH. Collagen Type III (COL3A1) was significantly upregulated by TGFβ1 in prostate stromal cells (PrSCs) and might be involved in DNM3OS function in myofibroblasts upon TGFβ1 stimulation. Upon TGFβ1 stimulation, COL3A1 protein was decreased by DNM3OS silencing. miR-29a and miR-29b could directly bind to the DNM3OS and COL3A1 3' untranslated region (UTR)s to negatively regulate their expression, and by serving as a competing endogenous RNAs (ceRNA), DNM3OS competed with COL3A1 for miR-29a/29b binding, therefore counteracting miR-29a/29b-mediated COL3A1 suppression. The effect of DNM3OS silencing on ECM components and TGFβ1 downstream signaling was similar to that of the TGFβ1 inhibitor SB431542. miR-361 could target DNM3OS and TGFβ1; DNM3OS competed for miR-361 binding to counteract miR-361-mediated TGFβ1 suppression. In conclusion, we identified DNM3OS as a specifically-upregulated lncRNA upon TGFβ1 stimulation in PrSCs; by serving as a ceRNA for the miR-29a/29b cluster and miR-361, DNM3OS eliminated miRNA-mediated suppression of COL3A1 and TGFβ1, thereby promoting TGFβ1-induced PrSC transformation into myofibroblasts.
Keywords: COL3A1; TGF-β1; benign prostatic hyperplasia (BPH); lncRNA DNM3OS; miR-29a/29b.
Conflict of interest statement
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
