

Hypoxia and EGF Stimulation Regulate VEGF Expression in Human Glioblastoma Multiforme (GBM) Cells by Differential Regulation of the PI3K/Rho-GTPase and MAPK Pathways
- PMID: 31698752
- PMCID: PMC6912653
- DOI: 10.3390/cells8111397
Hypoxia and EGF Stimulation Regulate VEGF Expression in Human Glioblastoma Multiforme (GBM) Cells by Differential Regulation of the PI3K/Rho-GTPase and MAPK Pathways
Abstract
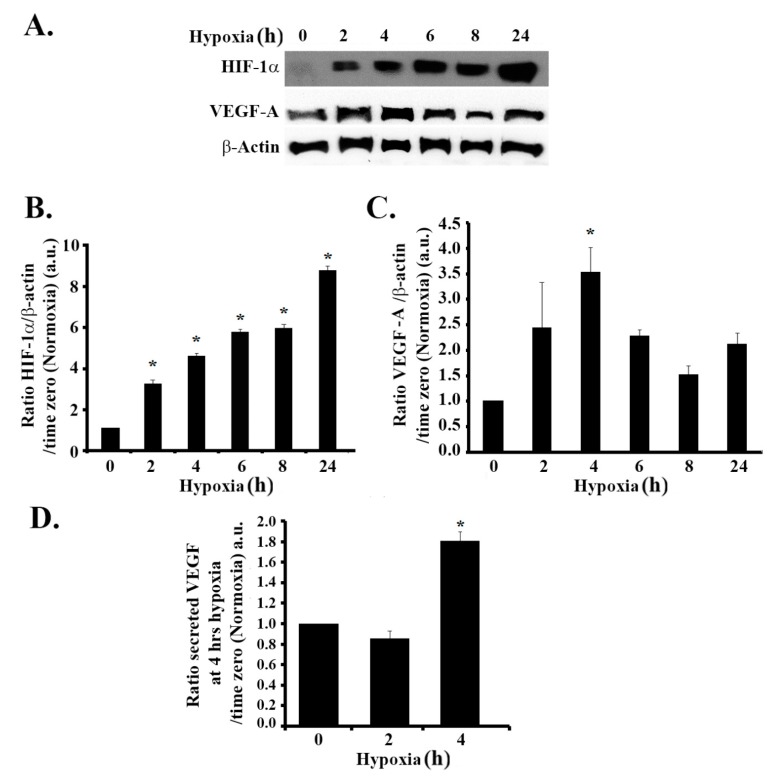
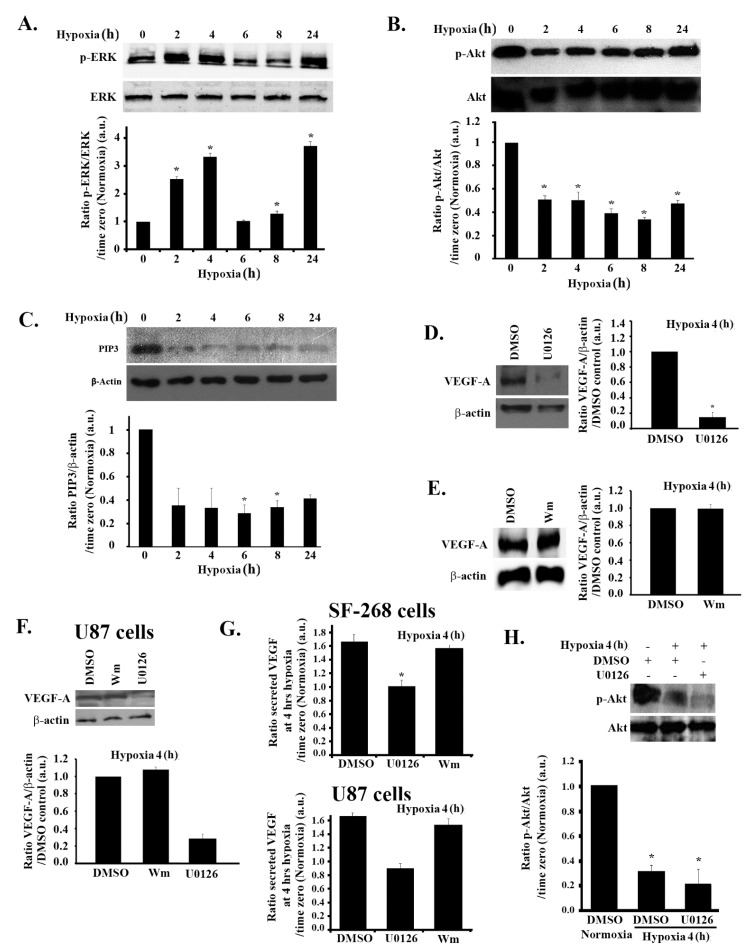
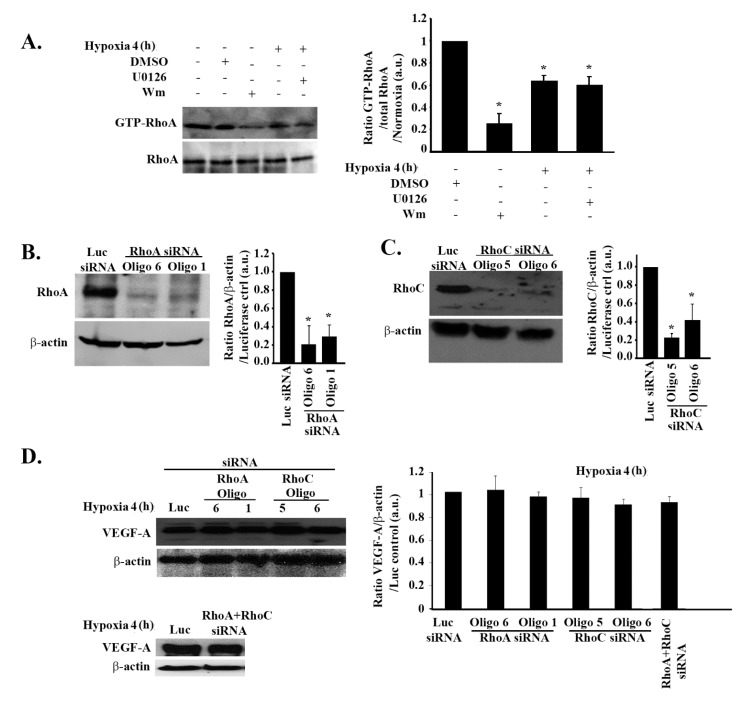
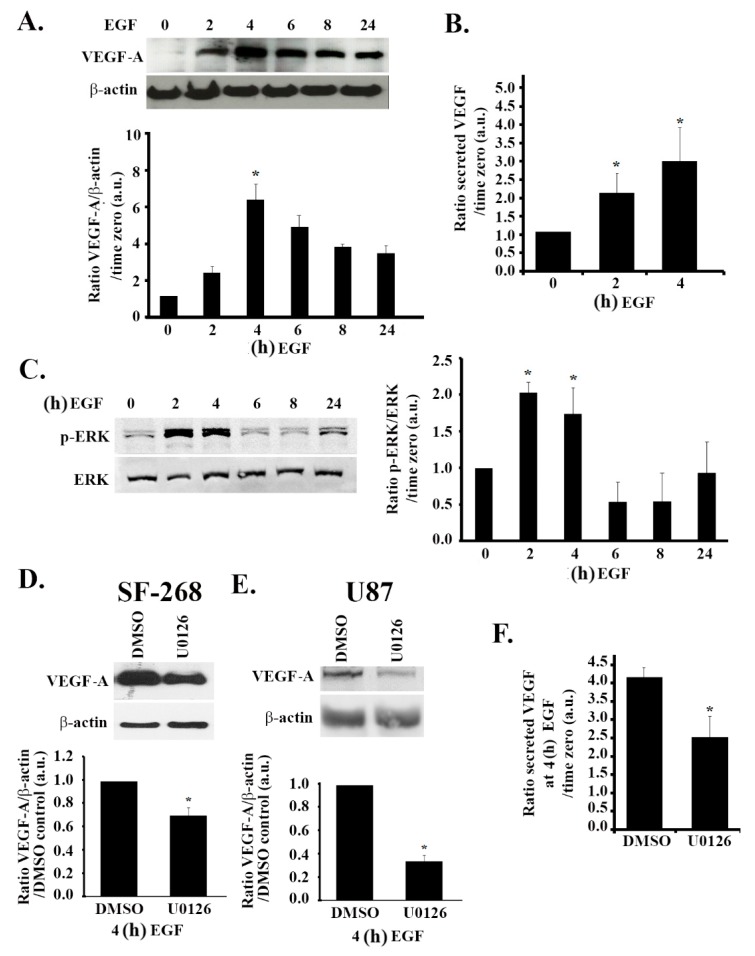
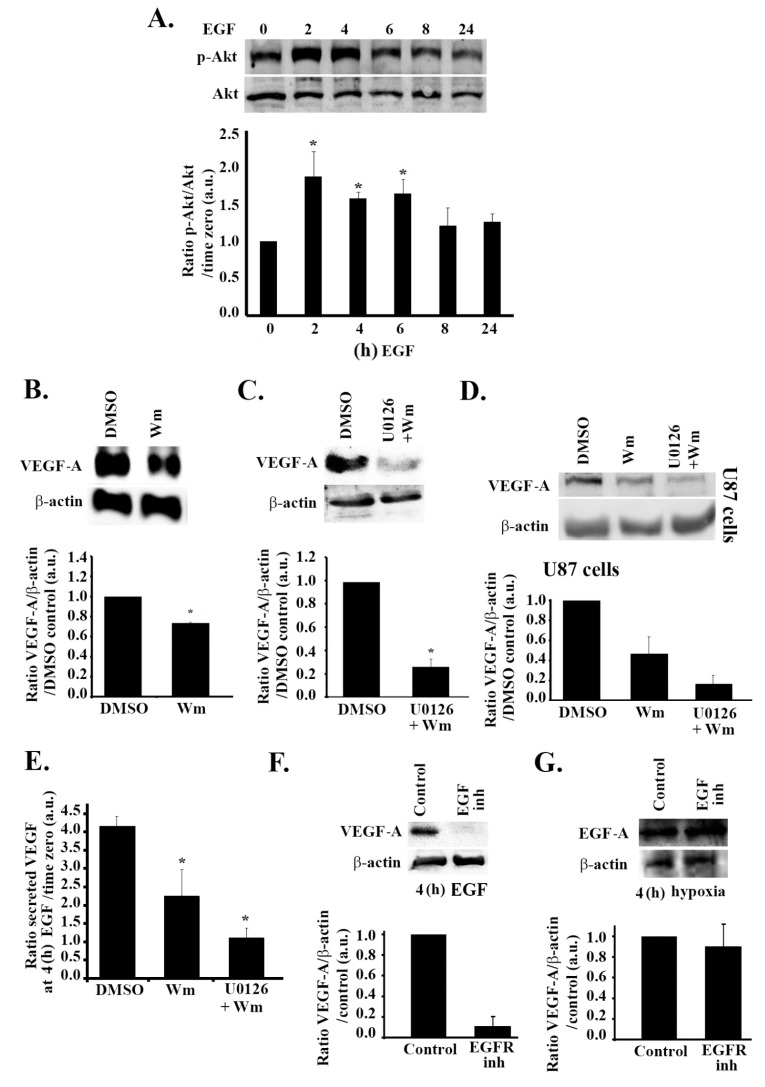
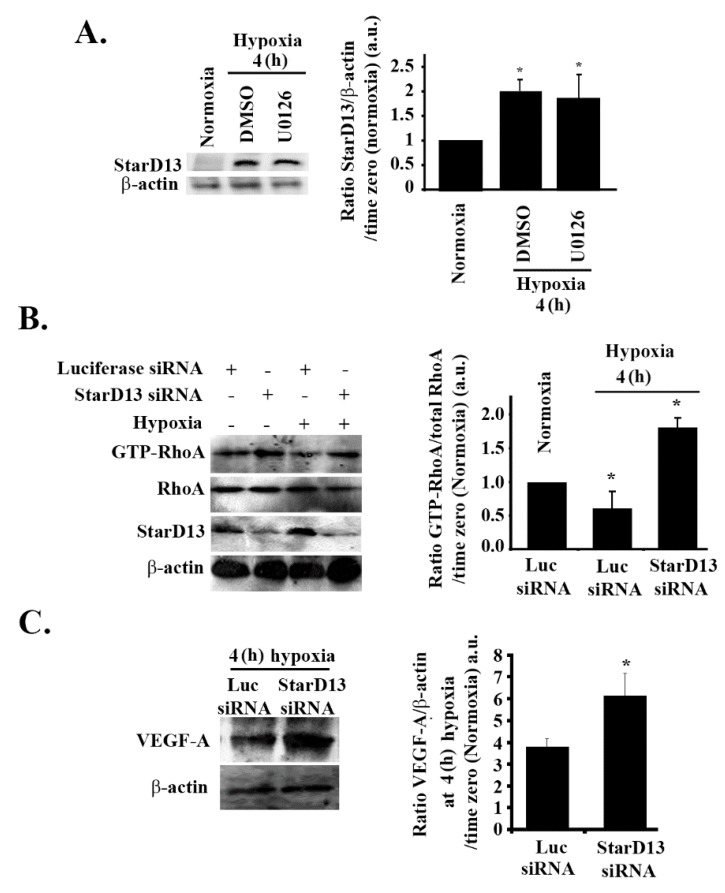
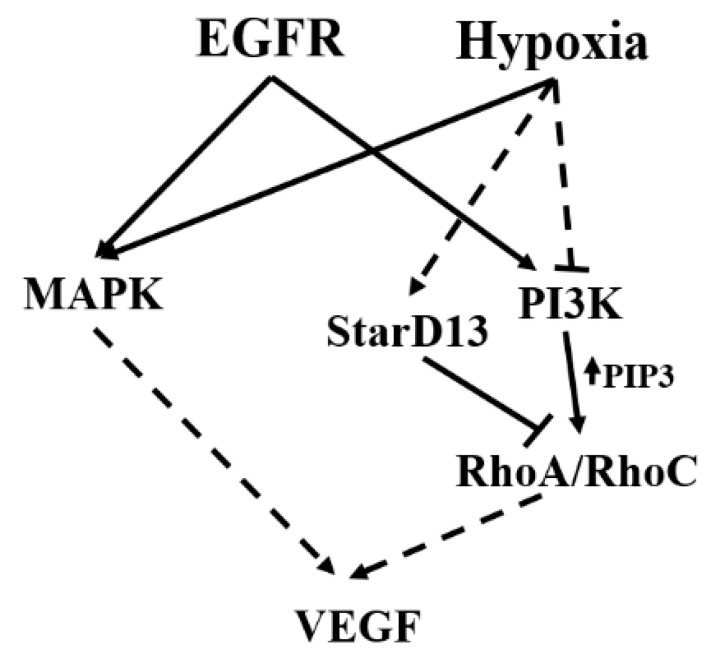
Glioblastoma multiforme (GBM) is one of the most common and deadly cancers of the central nervous system (CNS). It is characterized by the presence of hypoxic regions, especially in the core, leading to an increase in vascularity. This increased vascularization is driven by the expression of the major angiogenic inducer VEGF and the indirect angiogenic inducer Epidermal growth factor (EGF), which stimulates VEGF expression. In this study, we examine the regulation of VEGF by both hypoxia and the EGF signaling pathway. We also examine the involvement of pathways downstream from EGF signaling, including the mitogen-activated protein kinase/extracellular regulated kinase (MAPK/ERK) pathway and the Phosphatidylinositol-3-kinase/RhoA/C (PI3K/RhoA/C) pathway in this regulation. Our results show that VEGF expression and secretion levels increase following either hypoxia or EGF stimulation, with the two stimuli signaling in parallel. We also observed an increase in ERK and protein kinase B (Akt) phosphorylation, in response to EGF stimulation, with kinetics that correlated with the kinetics of the effect on VEGF. Using pharmacological inhibitors against ERK and PI3K and small interfering RNAs (siRNAs) against RhoA and RhoC, we found that both the ERK and the PI3K/RhoA/C pathways have to cooperate in order to lead to an increase in VEGF expression, downstream from EGF. In response to hypoxia, however, only ERK was involved in the regulation of VEGF. Hypoxia also led to a surprising decrease in the activation of PI3K and RhoA/C. Finally, the decrease in the activation of these Rho-GTPases was found to be mediated through a hypoxia-driven overexpression of the Rho-GTPase GTPase activating protein (GAP), StarD13. Therefore, while under normoxic conditions, EGF stimulates the activation of both the PI3K and the MAPK pathways and the induction of VEGF, in glioblastoma cells, hypoxic conditions lead to the suppression of the PI3K/RhoA/C pathway and an exclusive switch to the MAPK pathway.
Keywords: EGF; MAPK; PI3K; Rho-GTPases; VEGF; glioblastoma; hypoxia.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Vaupel P., Thews O., Kelleher D.K., Hoeckel M. Current status of knowledge and critical issues in tumor oxygenation. Results from 25 years research in tumor pathophysiology. Adv. Exp. Med. Biol. 1998;454:591–602. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
