

Rapid deacetylation of yeast Hsp70 mediates the cellular response to heat stress
- PMID: 31700027
- PMCID: PMC6838335
- DOI: 10.1038/s41598-019-52545-3
Rapid deacetylation of yeast Hsp70 mediates the cellular response to heat stress
Abstract
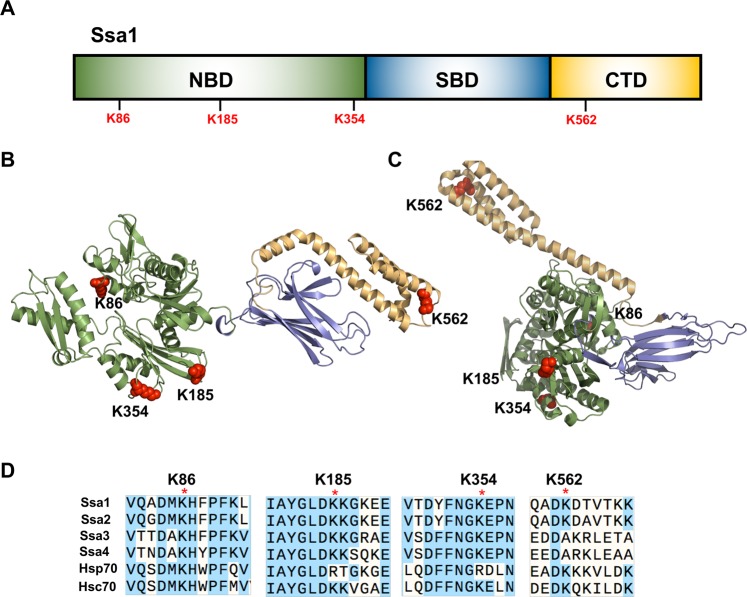
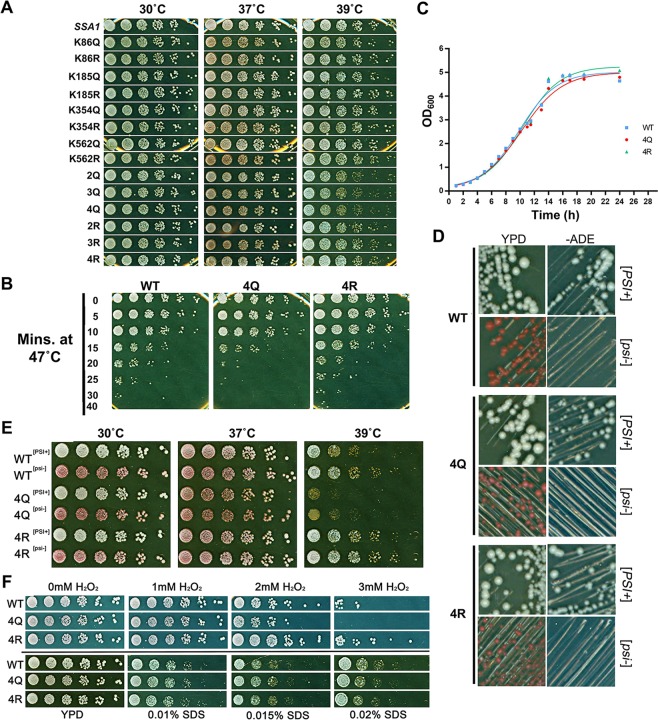
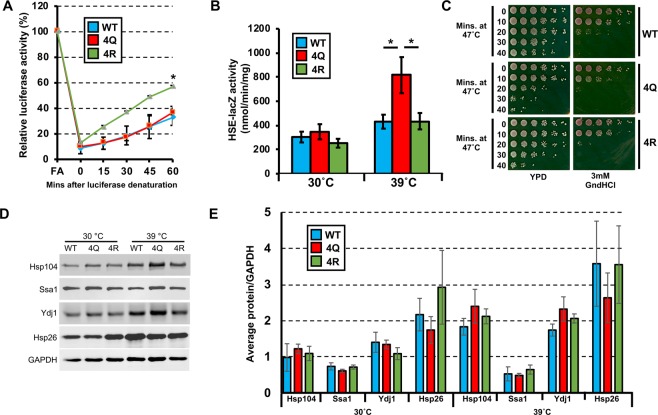
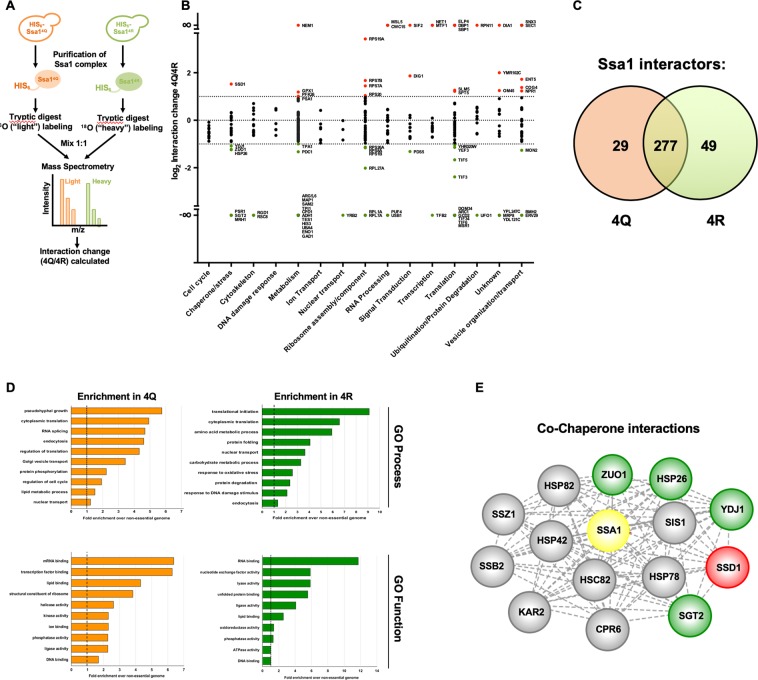
Hsp70 is a highly conserved molecular chaperone critical for the folding of new and denatured proteins. While traditional models state that cells respond to stress by upregulating inducible HSPs, this response is relatively slow and is limited by transcriptional and translational machinery. Recent studies have identified a number of post-translational modifications (PTMs) on Hsp70 that act to fine-tune its function. We utilized mass spectrometry to determine whether yeast Hsp70 (Ssa1) is differentially modified upon heat shock. We uncovered four lysine residues on Ssa1, K86, K185, K354 and K562 that are deacetylated in response to heat shock. Mutation of these sites cause a substantial remodeling of the Hsp70 interaction network of co-chaperone partners and client proteins while preserving essential chaperone function. Acetylation/deacetylation at these residues alter expression of other heat-shock induced chaperones as well as directly influencing Hsf1 activity. Taken together our data suggest that cells may have the ability to respond to heat stress quickly though Hsp70 deacetylation, followed by a slower, more traditional transcriptional response.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
