

Multiparameter Analysis Identifies Heterogeneity in Knee Osteoarthritis Synovial Responses
- PMID: 31702112
- PMCID: PMC7113107
- DOI: 10.1002/art.41161
Multiparameter Analysis Identifies Heterogeneity in Knee Osteoarthritis Synovial Responses
Abstract
Objective: Synovial membrane inflammation is common in osteoarthritis (OA) and increases cartilage injury. However, synovial fluid and histology studies suggest that OA inflammatory responses are not homogeneous. Greater understanding of these responses may provide new insights into OA disease mechanisms. We undertook this study to develop a novel multiparameter approach to phenotype synovial responses in knee OA.
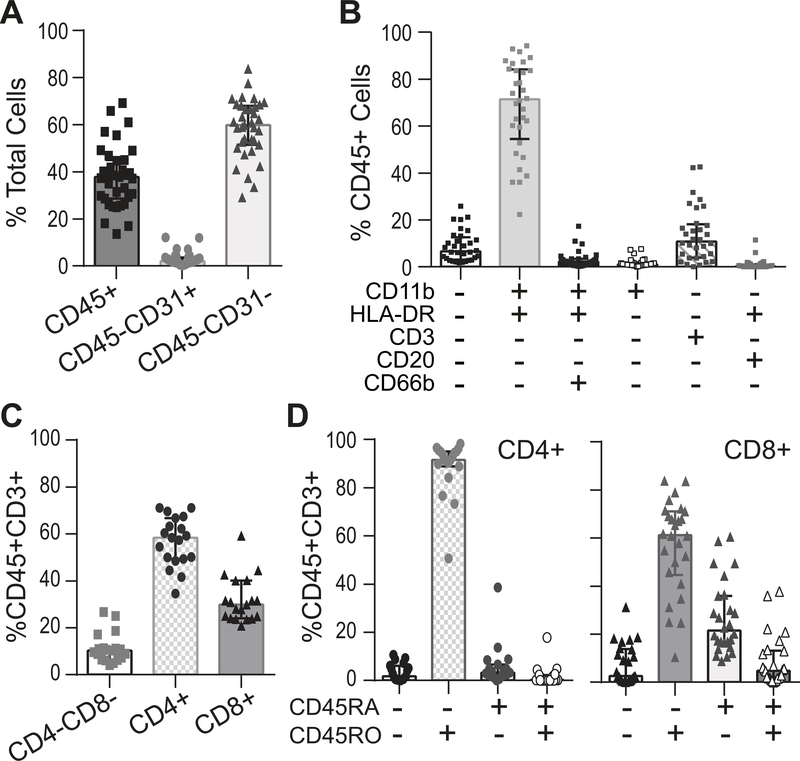
Methods: Cell composition and soluble protein production were measured by flow cytometry and multiplex enzyme-linked immunosorbent assay in synovium collected from OA patients undergoing knee replacement surgery (n = 35).
Results: Testing disaggregation conditions showed that aggressive digestion improved synovial cell yield and mesenchymal staining by flow cytometry, but it negatively impacted CD4+ T cell and CD56+ natural killer cell staining. Less aggressive digestion preserved these markers and showed highly variable T cell infiltration (range 0-43%; n = 32). Correlation analysis identified mesenchymal subpopulations associated with different nonmesenchymal populations, including macrophages and T cells (CD45+CD11b+HLA-DR+ myeloid cells with PDPN+CD73+CD90-CD34- mesenchymal cells [r = 0.65, P < 0.0001]; and CD45+CD3+ T cells with PDPN+CD73+CD90+CD34+ mesenchymal cells [r = 0.50, P = 0.003]). Interleukin-6 (IL-6) measured by flow cytometry strongly correlated with IL-6 released by ex vivo culture of synovial tissue (r = 0.59, P = 0.0012) and was highest in mesenchymal cells coexpressing CD90 and CD34. IL-6, IL-8, complement factor D, and IL-10 release correlated positively with tissue cellularity (P = 0.0042, P = 0.018, P = 0.0012, and P = 0.038, respectively). Additionally, increased CD8+ T cell numbers correlated with retinol binding protein 4 (P = 0.033). Finally, combining flow cytometry and multiplex data identified patient clusters with different types of inflammatory responses.
Conclusion: We used a novel approach to analyze OA synovium, identifying patient-specific inflammatory clusters. Our findings indicate that phenotyping synovial inflammation may provide new insights into OA patient heterogeneity and biomarker development.
© 2019, American College of Rheumatology.
Figures





References
-
- Beekhuizen M, Gierman LM, van Spil WE, Van Osch GJ, Huizinga TW, Saris DB, et al. An explorative study comparing levels of soluble mediators in control and osteoarthritic synovial fluid. Osteoarthritis Cartilage. 2013;21(7):918–22. - PubMed
-
- Knoop J, van der Leeden M, Thorstensson CA, Roorda LD, Lems WF, Knol DL, et al. Identification of phenotypes with different clinical outcomes in knee osteoarthritis: data from the Osteoarthritis Initiative. Arthritis Care Res (Hoboken). 2011;63(11):1535–42. - PubMed
-
- Oehler S, Neureiter D, Meyer-Scholten C, Aigner T. Subtyping of osteoarthritic synoviopathy. Clin Exp Rheumatol. 2002;20(5):633–40. - PubMed
-
- van Spil WE, Jansen NW, Bijlsma JW, Reijman M, DeGroot J, Welsing PM, et al. Clusters within a wide spectrum of biochemical markers for osteoarthritis: data from CHECK, a large cohort of individuals with very early symptomatic osteoarthritis. Osteoarthritis Cartilage. 2012;20(7):745–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
