

Cryo-EM Structure of the Human FLCN-FNIP2-Rag-Ragulator Complex
- PMID: 31704029
- PMCID: PMC7008705
- DOI: 10.1016/j.cell.2019.10.036
Cryo-EM Structure of the Human FLCN-FNIP2-Rag-Ragulator Complex
Abstract
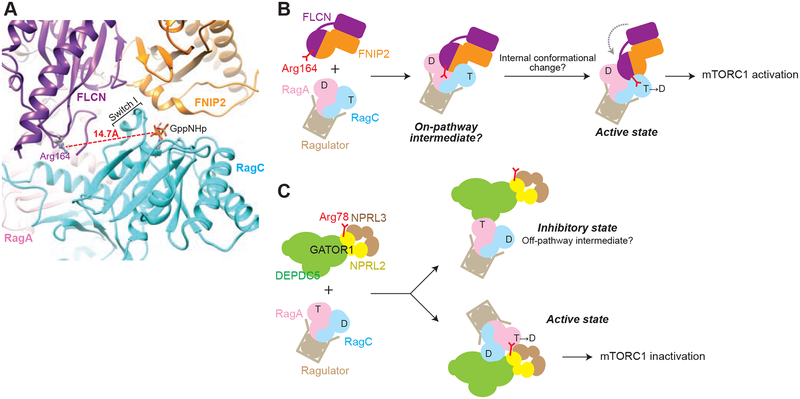
mTORC1 controls anabolic and catabolic processes in response to nutrients through the Rag GTPase heterodimer, which is regulated by multiple upstream protein complexes. One such regulator, FLCN-FNIP2, is a GTPase activating protein (GAP) for RagC/D, but despite its important role, how it activates the Rag GTPase heterodimer remains unknown. We used cryo-EM to determine the structure of FLCN-FNIP2 in a complex with the Rag GTPases and Ragulator. FLCN-FNIP2 adopts an extended conformation with two pairs of heterodimerized domains. The Longin domains heterodimerize and contact both nucleotide binding domains of the Rag heterodimer, while the DENN domains interact at the distal end of the structure. Biochemical analyses reveal a conserved arginine on FLCN as the catalytic arginine finger and lead us to interpret our structure as an on-pathway intermediate. These data reveal features of a GAP-GTPase interaction and the structure of a critical component of the nutrient-sensing mTORC1 pathway.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests
The authors declare no conflict of interest.
Figures
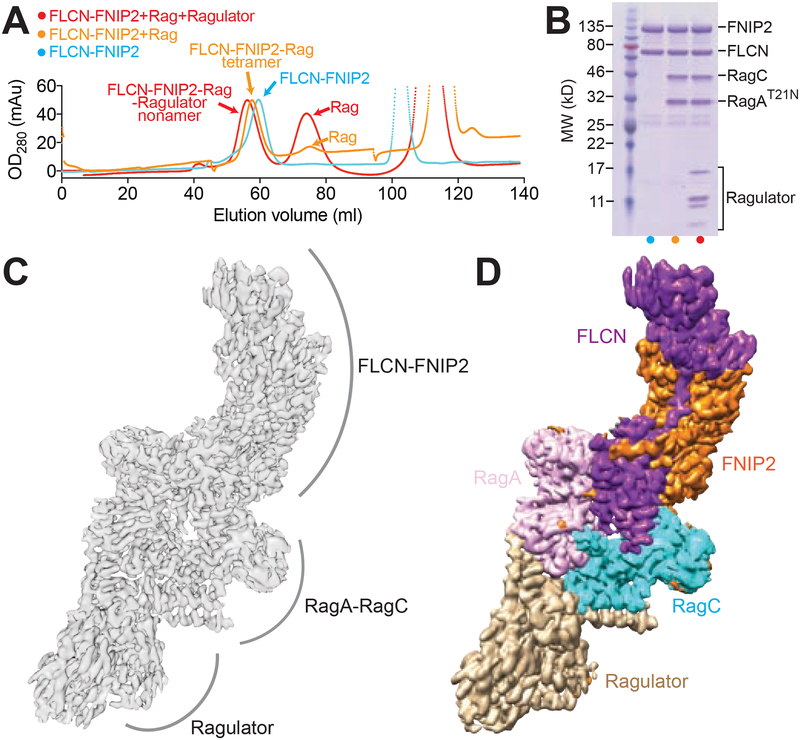
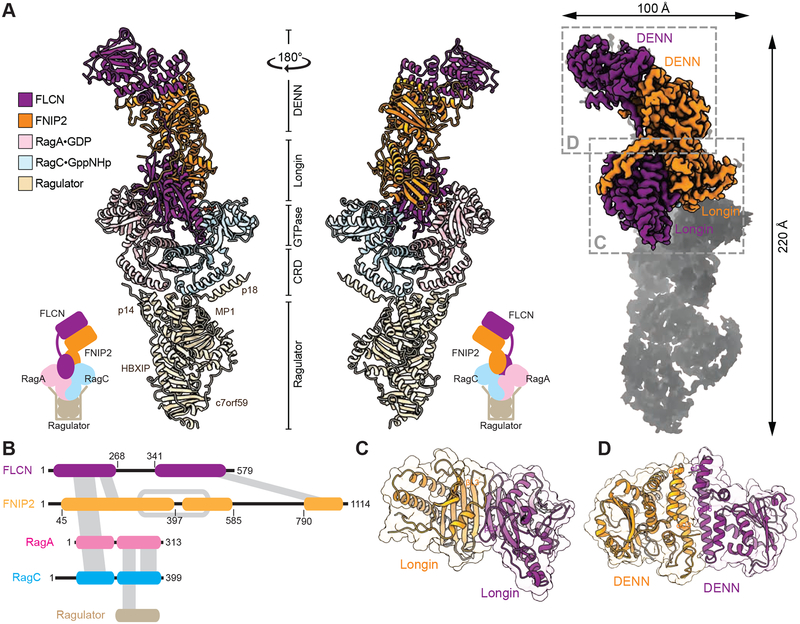
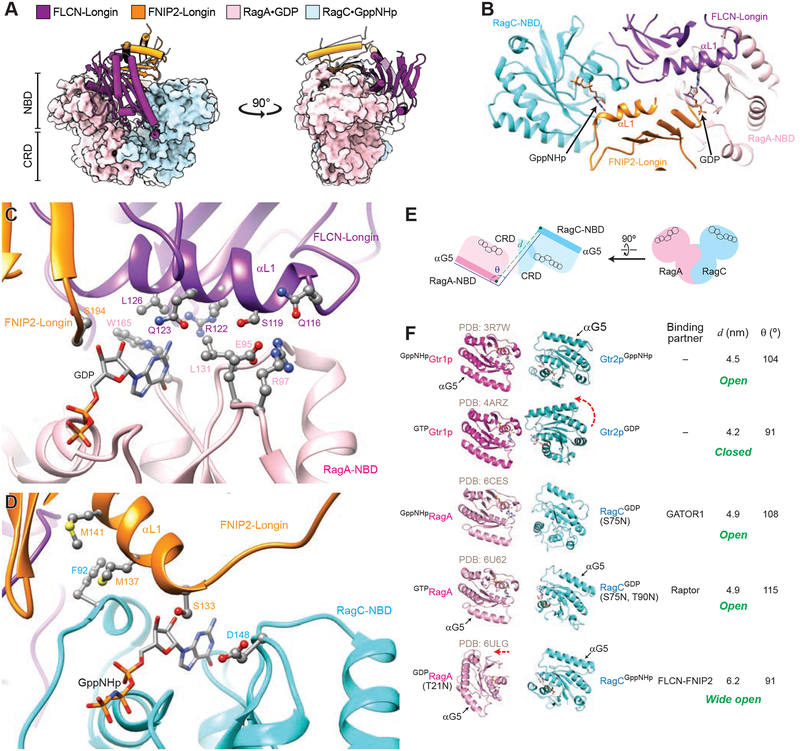
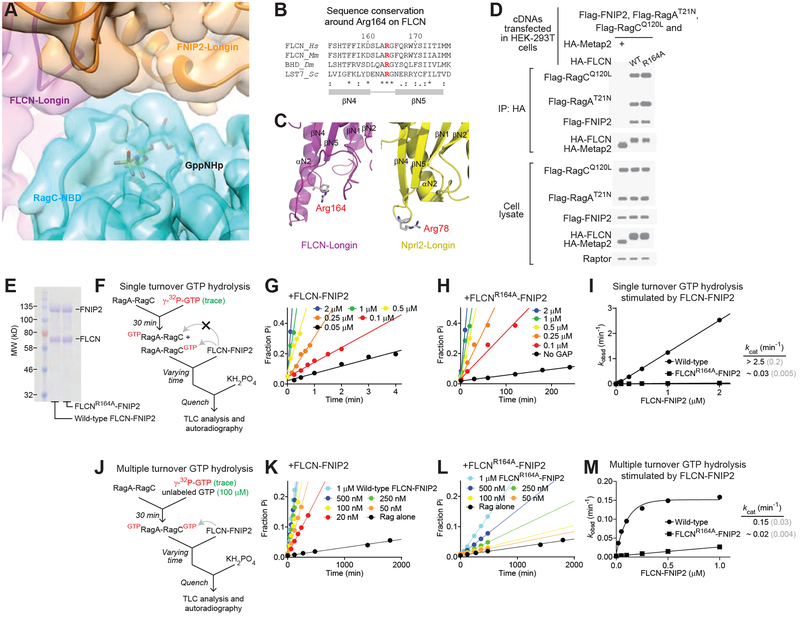
References
-
- Aylett CH, Sauer E, Imseng S, Boehringer D, Hall MN, Ban N, and Maier T (2016). Architecture of human mTOR complex 1. Science 351, 48–52. - PubMed
-
- Baba M, Hong SB, Sharma N, Warren MB, Nickerson ML, Iwamatsu A, Esposito D, Gillette WK, Hopkins RF 3rd, Hartley JL, et al. (2006). Folliculin encoded by the BHD gene interacts with a binding protein, FNIP1, and AMPK, and is involved in AMPK and mTOR signaling. Proc Natl Acad Sci USA 103, 15552–15557. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
