

Exfoliative toxin E, a new Staphylococcus aureus virulence factor with host-specific activity
- PMID: 31704997
- PMCID: PMC6841975
- DOI: 10.1038/s41598-019-52777-3
Exfoliative toxin E, a new Staphylococcus aureus virulence factor with host-specific activity
Erratum in
-
Author Correction: Exfoliative toxin E, a new Staphylococcus aureus virulence factor with host-specific activity.Sci Rep. 2020 Feb 7;10(1):2460. doi: 10.1038/s41598-020-59360-1. Sci Rep. 2020. PMID: 32034299 Free PMC article.
Abstract
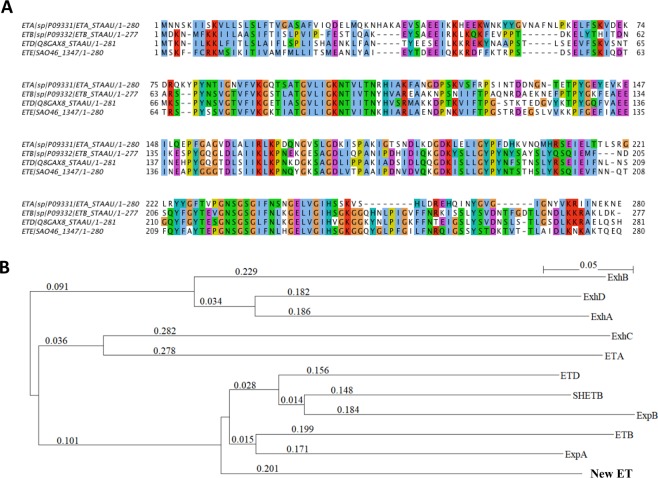
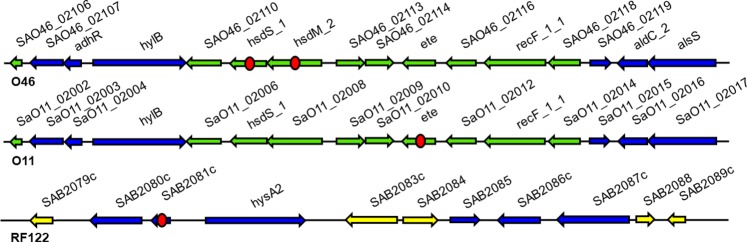
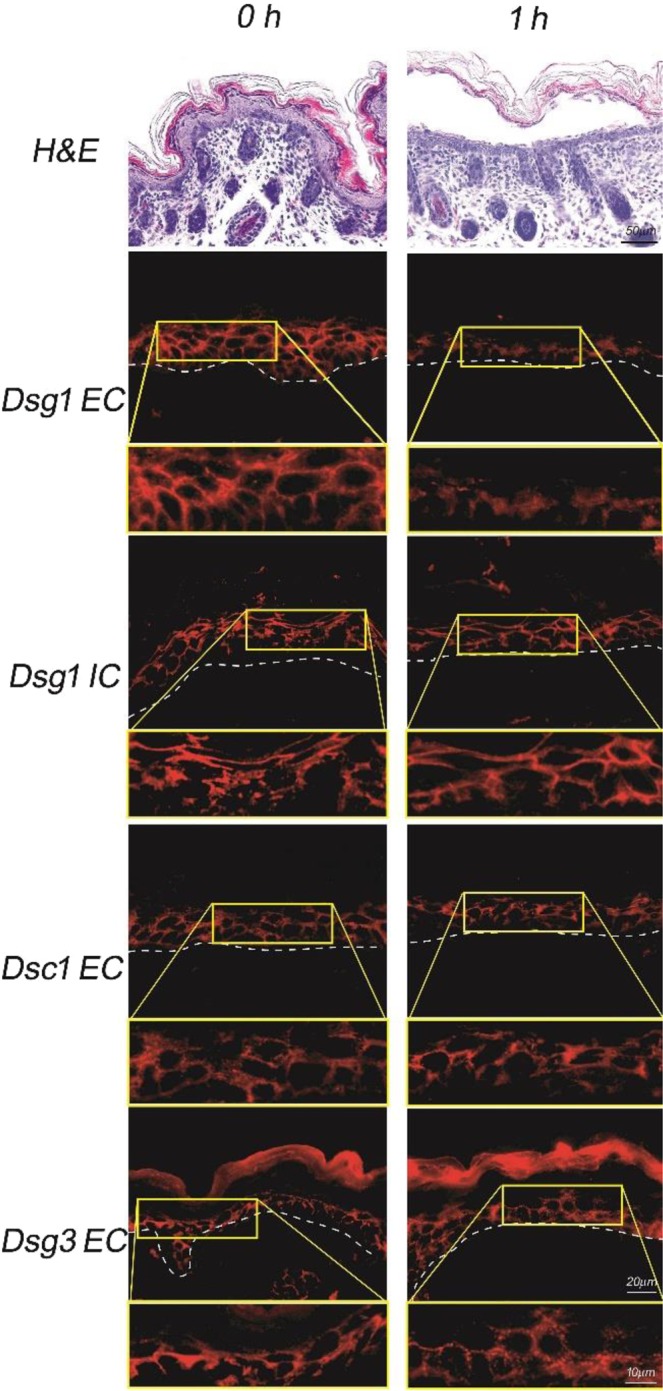
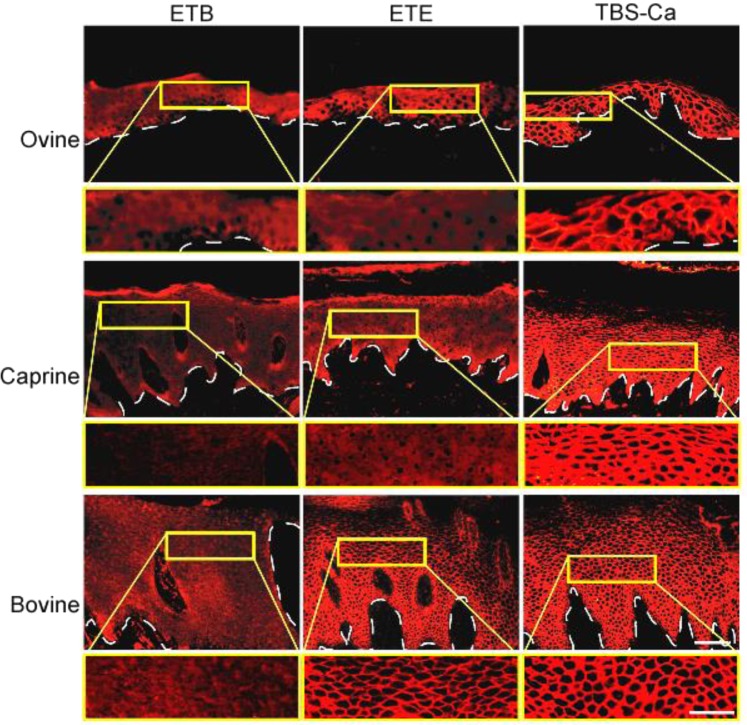
Exfoliative toxins (ETs) are secreted virulence factors produced by staphylococci. These serine proteases specifically cleave desmoglein 1 (Dsg1) in mammals and are key elements in staphylococcal skin infections. We recently identified a new et gene in S. aureus O46, a strain isolated from ovine mastitis. In the present study, we characterized the new et gene at a genetic level and the enzymatic activity of the deduced protein. The S. aureus O46 genome was re-assembled, annotated and compared with other publicly available S. aureus genomes. The deduced amino acid sequence of the new et gene shared 40%, 53% and 59% sequence identity to those of ETA, ETB and ETD, respectively. The new et gene shared the same genetic vicinity and was similar in other S. aureus strains bearing this gene. The recombinant enzyme of the new et gene caused skin exfoliation in vivo in neonatal mice. The new et-gene was thus named ete, encoding a new type (type E) of exfoliative toxin. We showed that ETE degraded the extracellular segments of Dsg1 in murine, ovine and caprine epidermis, as well as in ovine teat canal epithelia, but not that in bovine epidermis. We further showed that it directly hydrolyzed human and swine Dsg1 as well as murine Dsg1α and Dsg1β, but not canine Dsg1 or murine Dsg1γ. Molecular modeling revealed a correlation between the preferred orientation of ETE docking on its Dsg1 cleavage site and species-specific cleavage activity, suggesting that the docking step preceding cleavage accounts for the ETE species-specificity. This new virulence factor may contribute to the bacterial colonization on the stratified epithelia in certain ruminants with mastitis.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
