

The level of activity of the alternative lengthening of telomeres correlates with patient age in IDH-mutant ATRX-loss-of-expression anaplastic astrocytomas
- PMID: 31706351
- PMCID: PMC6842523
- DOI: 10.1186/s40478-019-0833-0
The level of activity of the alternative lengthening of telomeres correlates with patient age in IDH-mutant ATRX-loss-of-expression anaplastic astrocytomas
Abstract
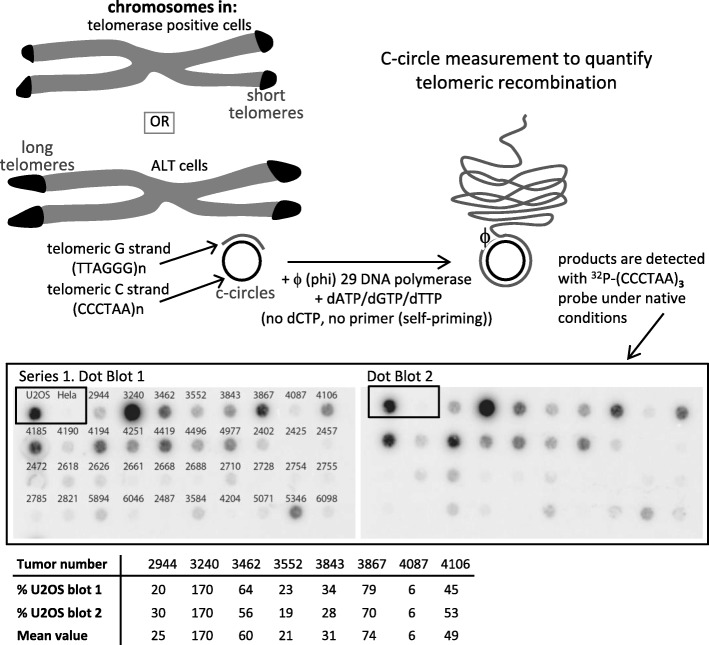
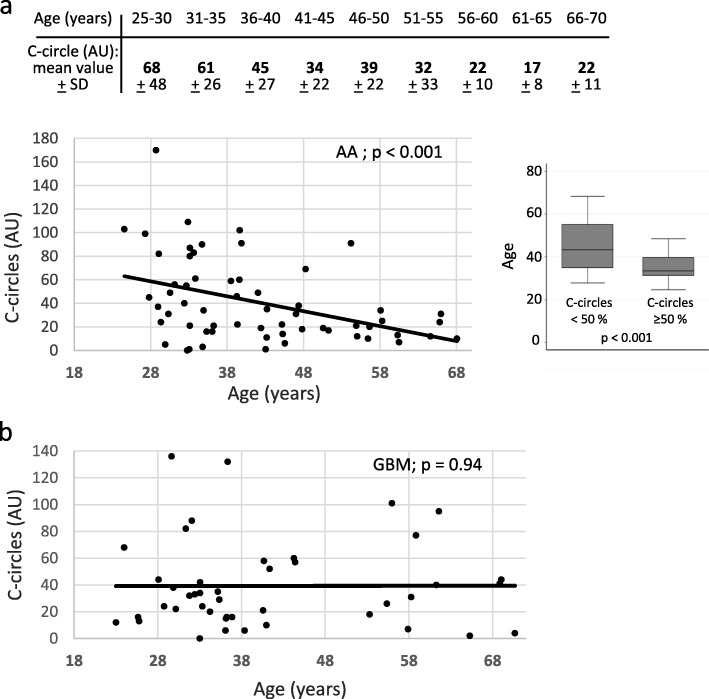
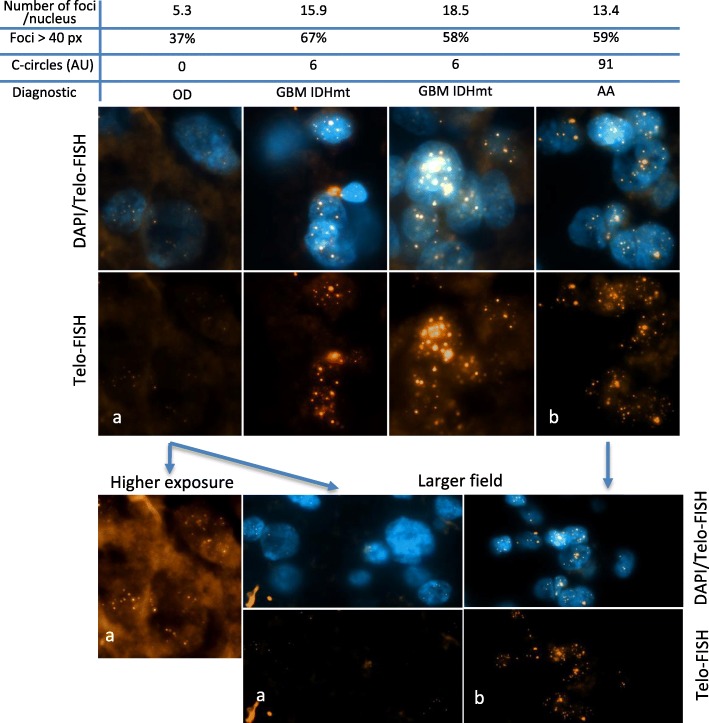
All cancer cells need to maintain functional telomeres to sustain continuous cell division and proliferation. In human diffuse gliomas, functional telomeres are maintained due either to reactivation of telomerase expression, the main pathway in most cancer types, or to activation of a mechanism called the alternative lengthening of telomeres (ALT). The presence of IDH1/2 mutations (IDH-mutant) together with loss of ATRX expression (ATRX-lost) are frequently associated with ALT in diffuse gliomas. However, detection of ALT, and a fortiori its quantification, are rarely, if ever, measured in neuropathology laboratories. We measured the level of ALT activity using the previously described quantitative "C-circle" assay and analyzed it in a well characterized cohort of 104 IDH-mutant and ATRX-lost adult diffuse gliomas. We report that in IDH-mutant ATRX-lost anaplastic astrocytomas, the intensity of ALT was inversely correlated with age (p < 0.001), the younger the patient, the higher the intensity of ALT. Strikingly, glioblastomas having progressed from anaplastic astrocytomas did not exhibit this correlation. ALT activity level in the tumor did not depend on telomere length in healthy tissue cells from the same patient. In summary, we have uncovered the existence, in anaplastic astrocytomas but not in glioblastomas with the same IDH and ATRX mutations, of a correlation between patient age and the level of activity of ALT, a telomerase-independent pathway of telomere maintenance.
Keywords: ATRX loss of expression; Alternative lengthening of telomeres; Anaplastic astrocytoma; IDH1/2 mutations; Secondary glioblastoma.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
