

Microbial Metabolite Signaling Is Required for Systemic Iron Homeostasis
- PMID: 31708445
- PMCID: PMC6949377
- DOI: 10.1016/j.cmet.2019.10.005
Microbial Metabolite Signaling Is Required for Systemic Iron Homeostasis
Abstract
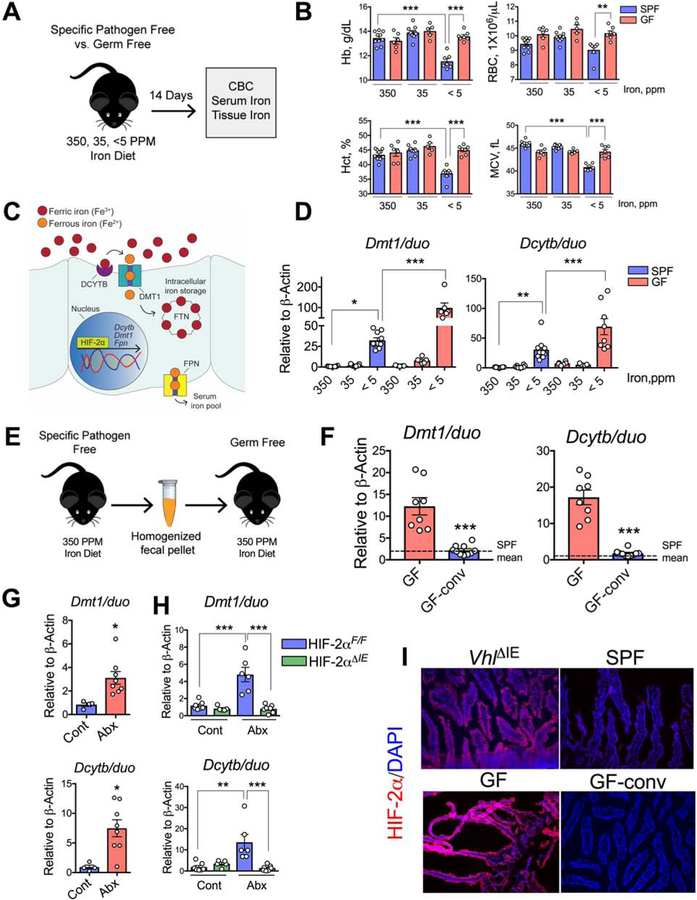
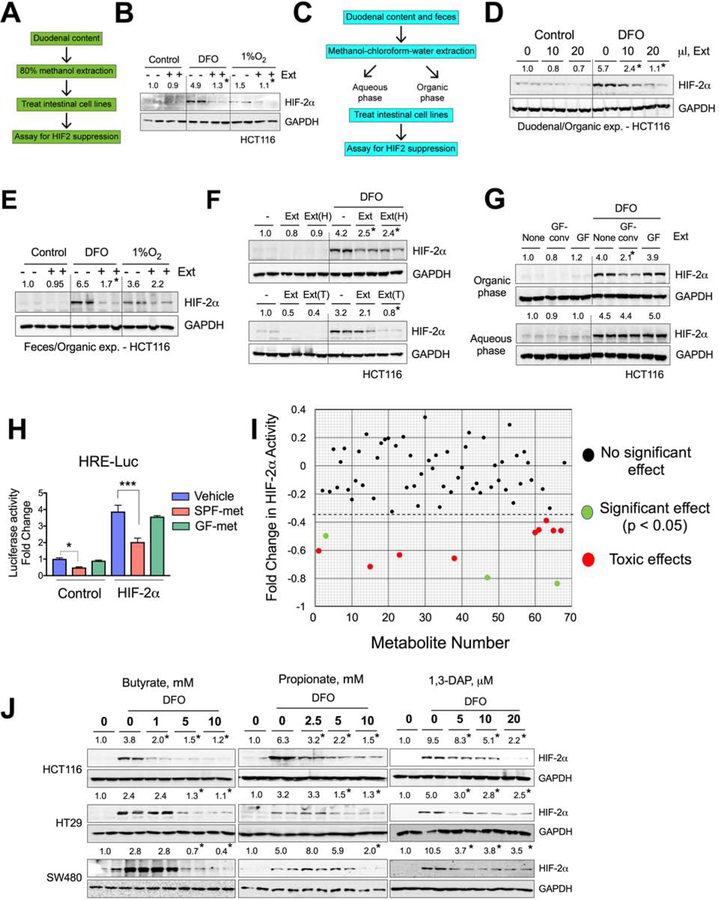
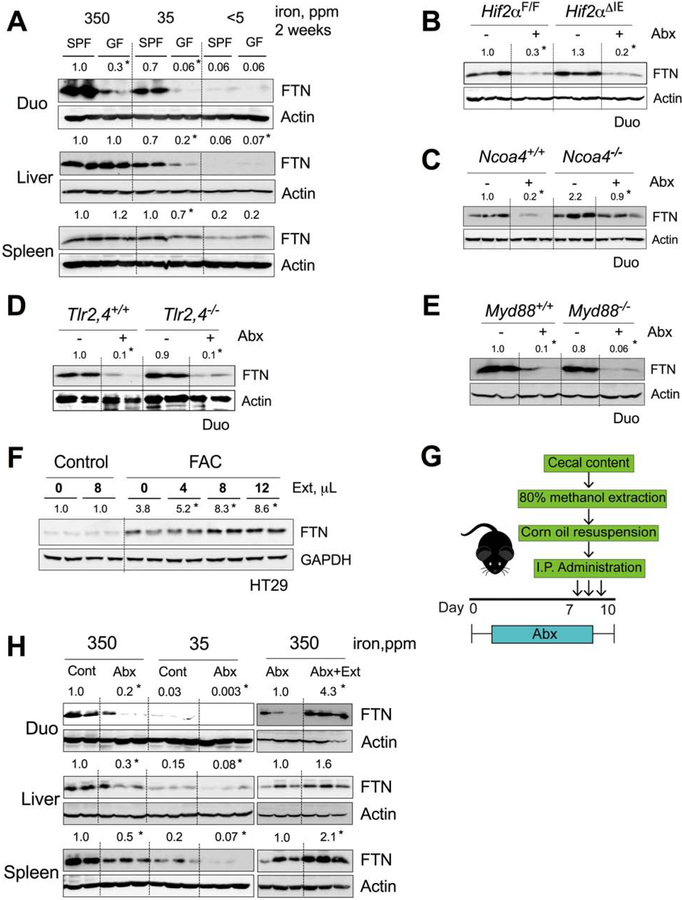
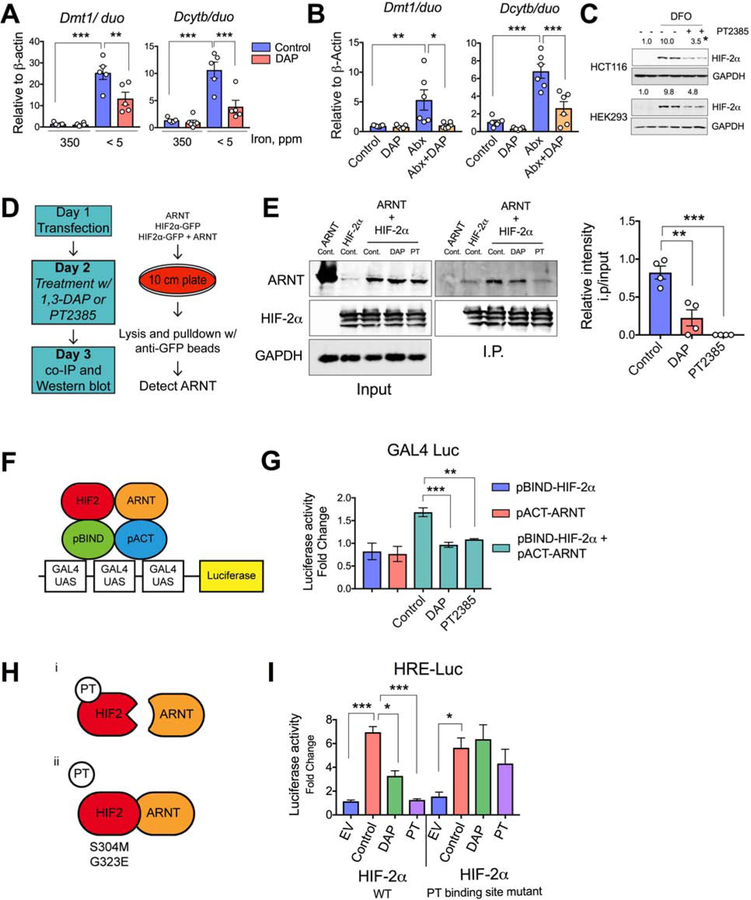
Iron is a central micronutrient needed by all living organisms. Competition for iron in the intestinal tract is essential for the maintenance of indigenous microbial populations and for host health. How symbiotic relationships between hosts and native microbes persist during times of iron limitation is unclear. Here, we demonstrate that indigenous bacteria possess an iron-dependent mechanism that inhibits host iron transport and storage. Using a high-throughput screen of microbial metabolites, we found that gut microbiota produce metabolites that suppress hypoxia-inducible factor 2α (HIF-2α) a master transcription factor of intestinal iron absorption and increase the iron-storage protein ferritin, resulting in decreased intestinal iron absorption by the host. We identified 1,3-diaminopropane (DAP) and reuterin as inhibitors of HIF-2α via inhibition of heterodimerization. DAP and reuterin effectively ameliorated systemic iron overload. This work provides evidence of intestine-microbiota metabolic crosstalk that is essential for systemic iron homeostasis.
Keywords: EPAS1; HIF; HIF-2a; anemia; ferritin; hemochromatosis; hypoxia; hypoxia-inducible factor; iron; metabolites; microbiota.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
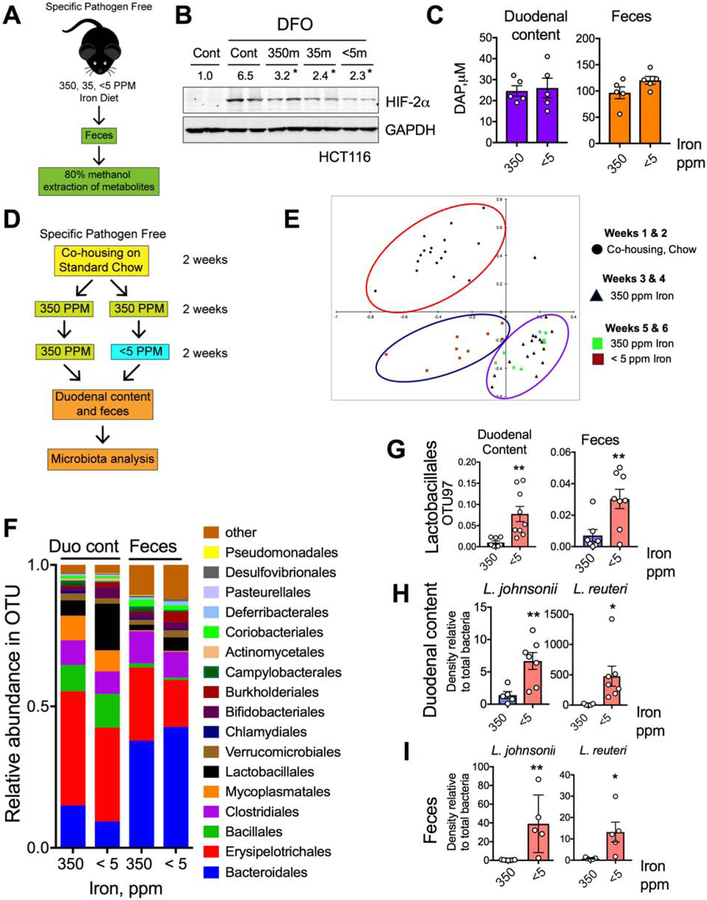
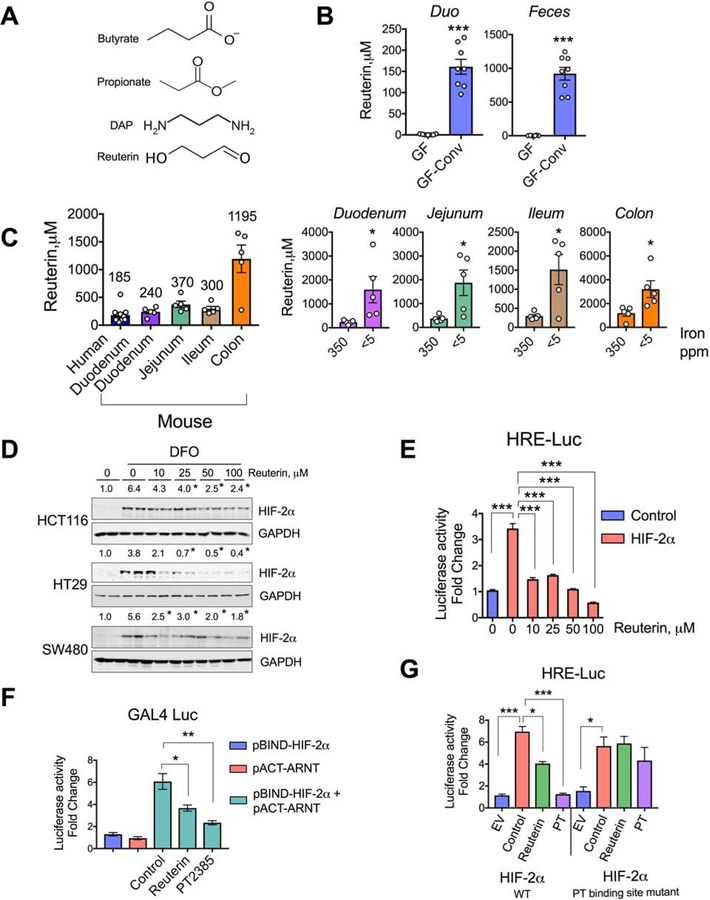
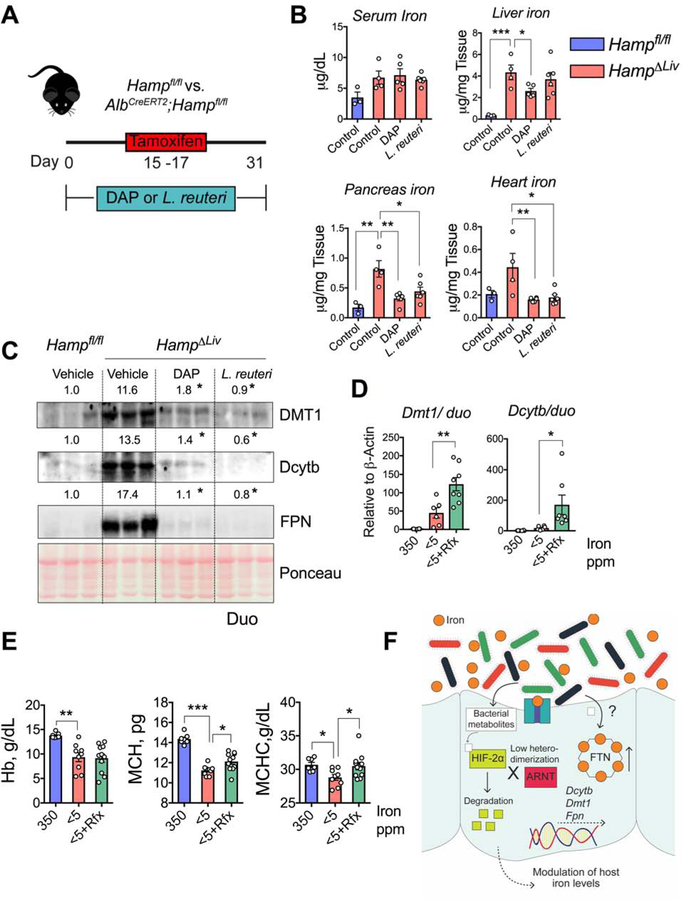
Comment in
-
Iron Deficiency Causes Wrinkles in Host-Commensal Relationships.Cell Metab. 2020 Jan 7;31(1):8-10. doi: 10.1016/j.cmet.2019.12.003. Cell Metab. 2020. PMID: 31951569
References
-
- Adams PC, and Barton JC (2007). Haemochromatosis. Lancet 370, 1855–1860. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
