

Fatty Liver Due to Increased de novo Lipogenesis: Alterations in the Hepatic Peroxisomal Proteome
- PMID: 31709254
- PMCID: PMC6823594
- DOI: 10.3389/fcell.2019.00248
Fatty Liver Due to Increased de novo Lipogenesis: Alterations in the Hepatic Peroxisomal Proteome
Abstract
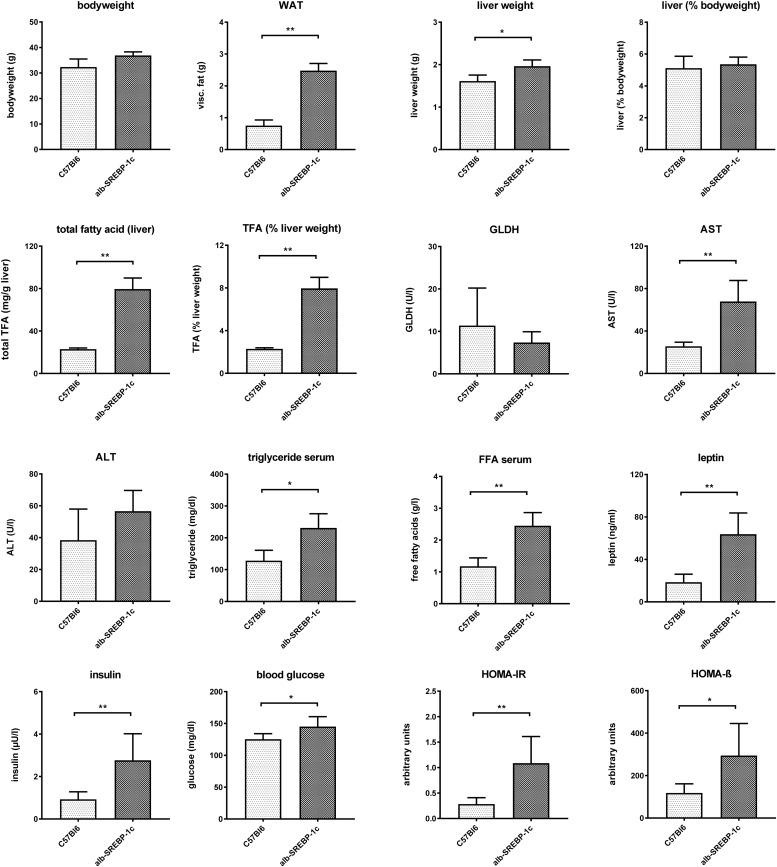
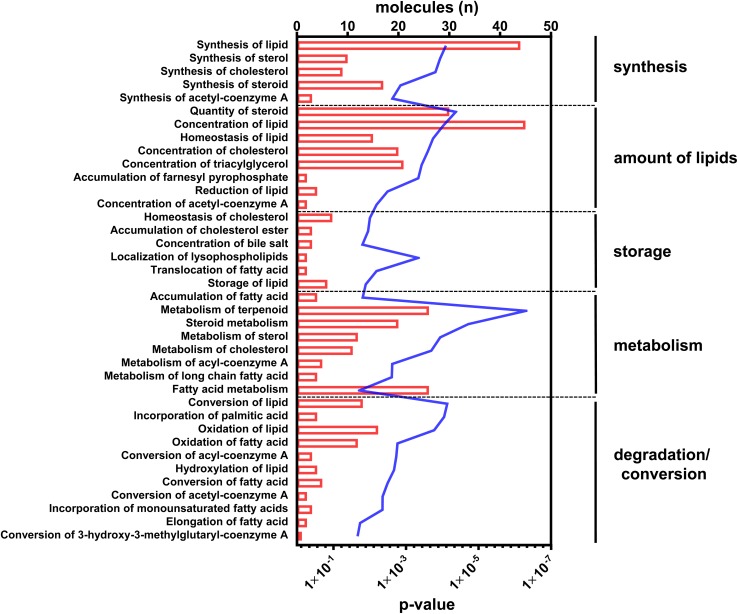
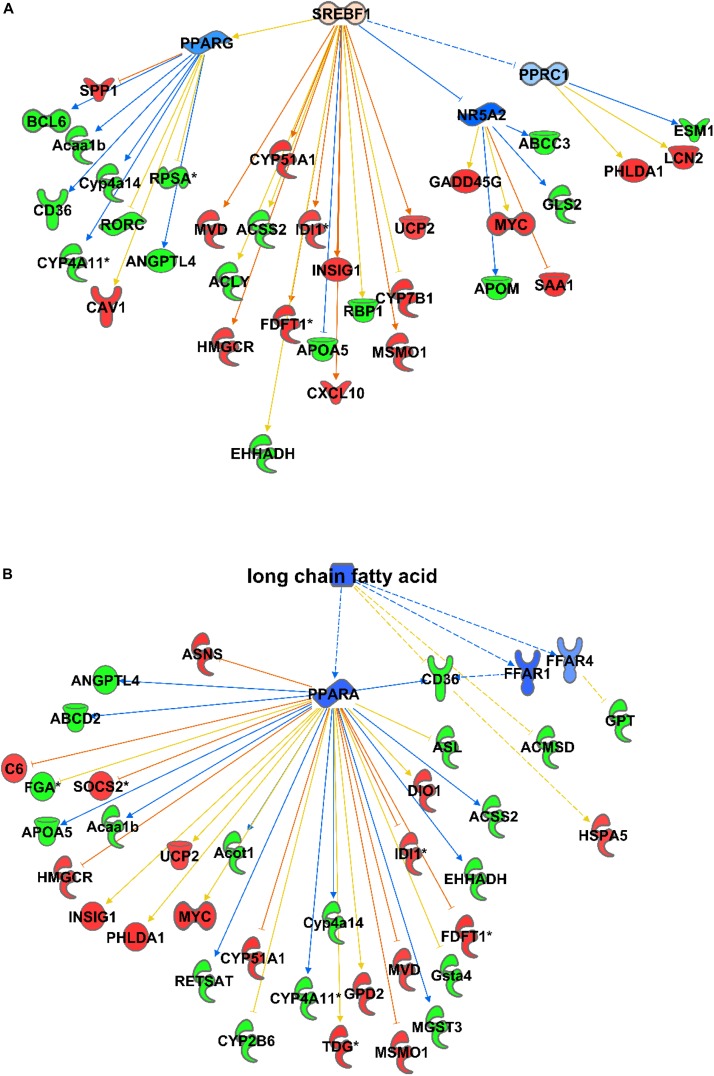
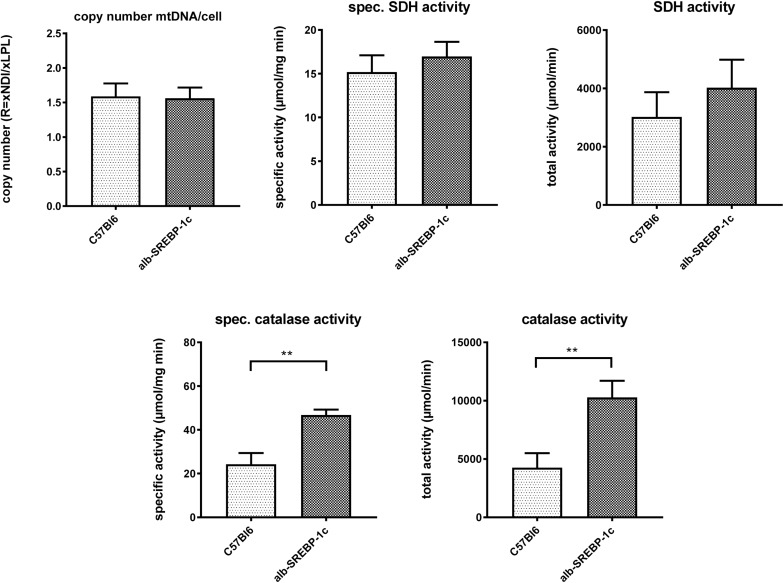
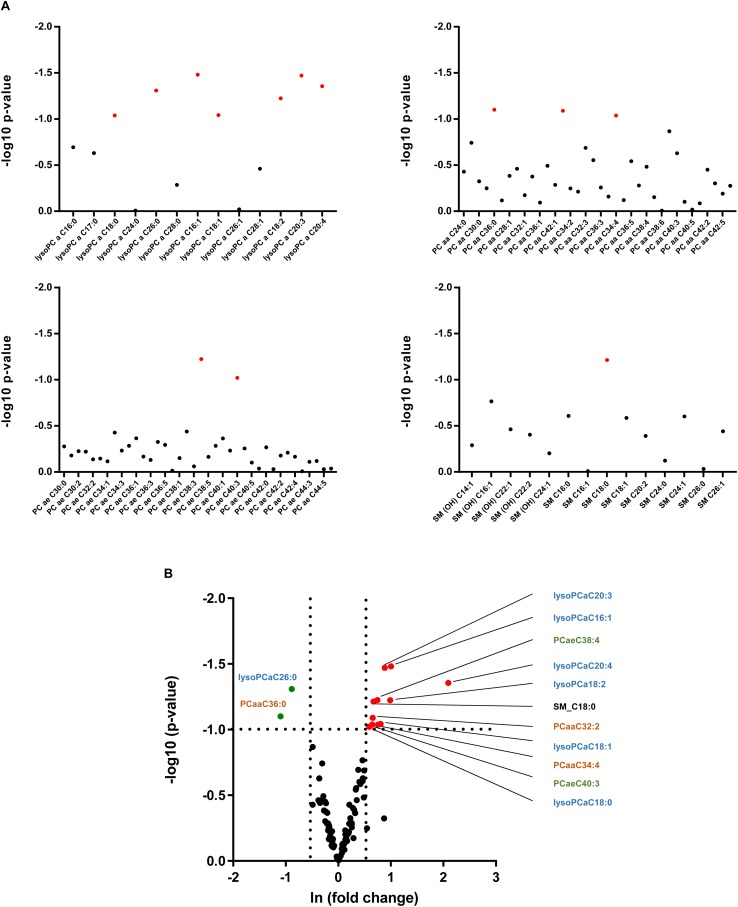
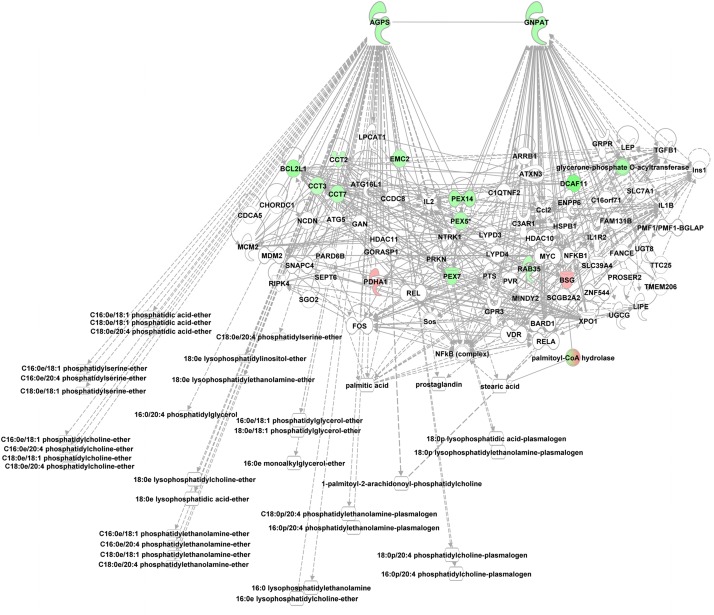
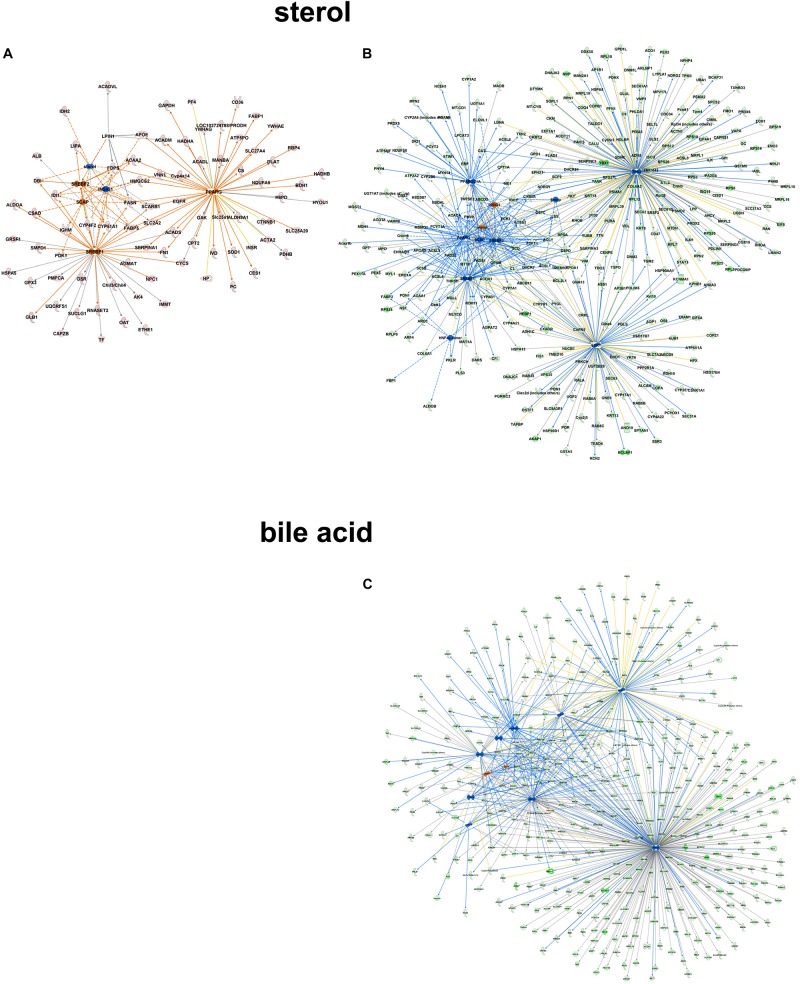
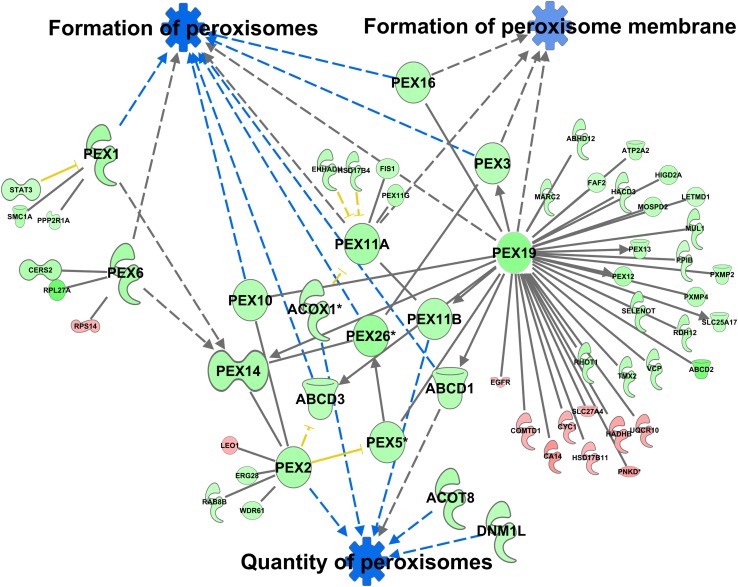
In non-alcoholic fatty liver disease (NAFLD) caused by ectopic lipid accumulation, lipotoxicity is a crucial molecular risk factor. Mechanisms to eliminate lipid overflow can prevent the liver from functional complications. This may involve increased secretion of lipids or metabolic adaptation to ß-oxidation in lipid-degrading organelles such as mitochondria and peroxisomes. In addition to dietary factors, increased plasma fatty acid levels may be due to increased triglyceride synthesis, lipolysis, as well as de novo lipid synthesis (DNL) in the liver. In the present study, we investigated the impact of fatty liver caused by elevated DNL, in a transgenic mouse model with liver-specific overexpression of human sterol regulatory element-binding protein-1c (alb-SREBP-1c), on hepatic gene expression, on plasma lipids and especially on the proteome of peroxisomes by omics analyses, and we interpreted the results with knowledge-based analyses. In summary, the increased hepatic DNL is accompanied by marginal gene expression changes but massive changes in peroxisomal proteome. Furthermore, plasma phosphatidylcholine (PC) as well as lysoPC species were altered. Based on these observations, it can be speculated that the plasticity of organelles and their functionality may be directly affected by lipid overflow.
Keywords: DNL; NAFLD; SREBP-1c; fatty liver; label-free proteomic profiling; lipidomics; peroxisomes; transcriptomics.
Copyright © 2019 Knebel, Fahlbusch, Dille, Wahlers, Hartwig, Jacob, Kettel, Schiller, Herebian, Koellmer, Lehr, Müller-Wieland and Kotzka.
Figures


References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous