

TBK1-mediated phosphorylation of LC3C and GABARAP-L2 controls autophagosome shedding by ATG4 protease
- PMID: 31709703
- PMCID: PMC6945063
- DOI: 10.15252/embr.201948317
TBK1-mediated phosphorylation of LC3C and GABARAP-L2 controls autophagosome shedding by ATG4 protease
Abstract
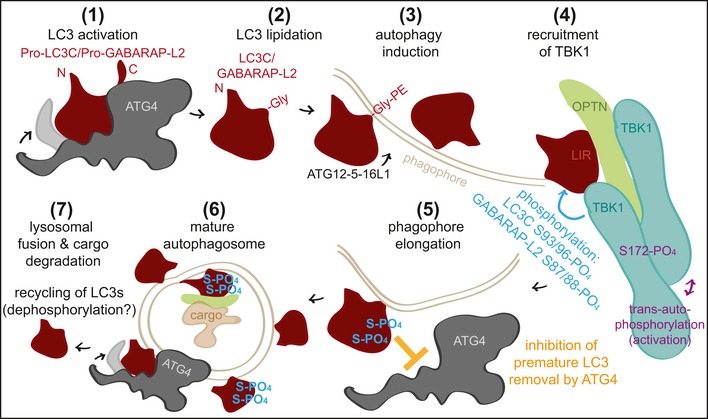
Autophagy is a highly conserved catabolic process through which defective or otherwise harmful cellular components are targeted for degradation via the lysosomal route. Regulatory pathways, involving post-translational modifications such as phosphorylation, play a critical role in controlling this tightly orchestrated process. Here, we demonstrate that TBK1 regulates autophagy by phosphorylating autophagy modifiers LC3C and GABARAP-L2 on surface-exposed serine residues (LC3C S93 and S96; GABARAP-L2 S87 and S88). This phosphorylation event impedes their binding to the processing enzyme ATG4 by destabilizing the complex. Phosphorylated LC3C/GABARAP-L2 cannot be removed from liposomes by ATG4 and are thus protected from ATG4-mediated premature removal from nascent autophagosomes. This ensures a steady coat of lipidated LC3C/GABARAP-L2 throughout the early steps in autophagosome formation and aids in maintaining a unidirectional flow of the autophagosome to the lysosome. Taken together, we present a new regulatory mechanism of autophagy, which influences the conjugation and de-conjugation of LC3C and GABARAP-L2 to autophagosomes by TBK1-mediated phosphorylation.
Keywords: ATG4; ATG8; TBK1; autophagy; phosphorylation.
© 2019 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
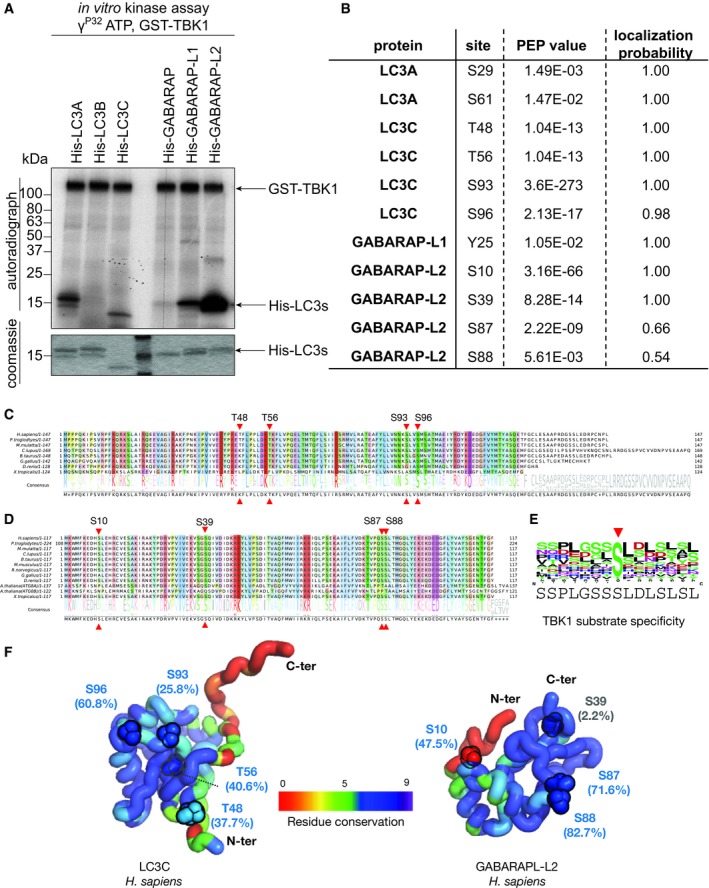
- A
Coomassie stain and autoradiography of SDS–PAGE after an in vitro kinase assay with GST‐TBK1 and His‐LC3 family proteins as substrates. TBK1 phosphorylates LC3A, LC3C, GABARAP‐L1, and GABARAP‐L2 in vitro.
- B
Identification of phosphosites by mass spectrometry following an in vitro TBK1 kinase assay.
- C, D
Alignments showing selected orthologs of human LC3C (C) and human GABARAP‐L2 (D) highlighting the relative conservation of phosphosites (red triangles). The position of phosphosites identified within these proteins is labeled on top of the alignment along with the consensus sequence and normalized residue‐conservation frequency at the bottom. Complete alignments including all the orthologs from higher eukaryotes (jawed vertebrates, Gnathostomata) for LC3C (n = 207) and GABARAP‐L2 (n = 254) are provided in source files.
- E
Sequence motif characteristic of TBK1 substrates containing the central phosphoserine position (red arrow) was computed by aligning 15‐residue sequence fragments from 22 experimentally verified substrates of TBK1 from KinaseNET (a human protein kinase knowledge database:
http://www.kinasenet.ca ). - F
3D structures of full‐length human LC3C and GABARAP‐L2 highlighting sequence conservation (rainbow‐colored side chains) and solvent accessibility (%) of the identified phosphosites. Conservation scores (0–9 scale) are obtained from full set of orthologous protein alignments using ConSurf 57.
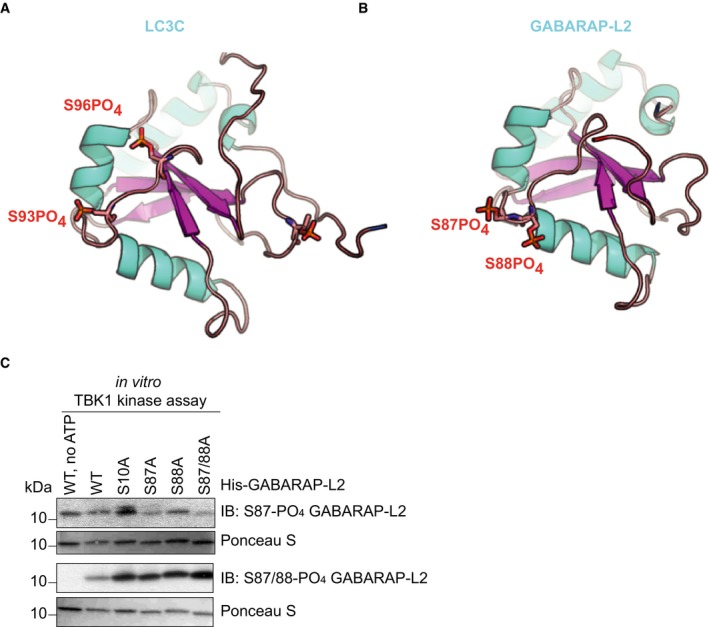
Structure of LC3C8–125 (PDB: 3WAM) with modeled phosphate groups (red sticks) at S93 and S96.
Topologically equivalent positions in GABARAP‐L2 (PDB: 4CO7), S87 and S88, are also phosphorylated (red sticks) by TBK1. Phosphorylation sites are on the opposite face of the LIR binding pocket of LC3 proteins.
SDS–PAGE and Western blot of in vitro TBK1 kinase assay with His‐GABARAP‐L2 WT or mutants as substrates to test the respective phospho‐GABARAP‐L2 antibodies for their specificity.
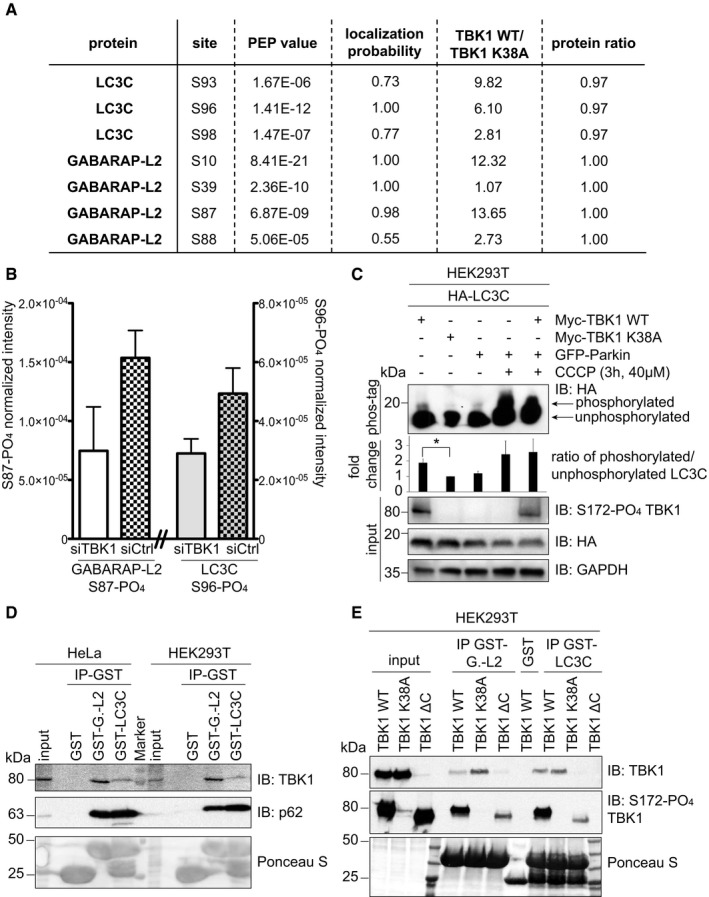
- A, B
Identification of phosphosites by mass spectrometry following GFP‐LC3C or GFP‐GABARAP‐L2 immunoprecipitation. (A) TBK1 WT was overexpressed in heavy‐ and TBK1 kinase‐dead K38A was overexpressed in light‐labeled SILAC HEK293T cells. (B) HEK293T cells were treated with control or TBK1 siRNA and CCCP (3 h, 40 μM). Phosphosite intensities were normalized to total protein intensity. Data are presented as mean ± SD, n = 3 biological replicates.
- C
SDS–PAGE and Western blot of Phos‐tag™ gel with HEK293T cell lysates. Cells were transfected with HA‐LC3C, TBK1 WT or K38A, and GFP‐Parkin and left untreated or treated with CCCP (3 h, 40 μM) to induce mitophagy. The ratio of phosphorylated to unphosphorylated LC3C was quantified. Data are presented as mean ± SD, n = 3 biological replicates, *P < 0.05, as analyzed by Student's t‐test.
- D
SDS–PAGE and Western blot of HEK293T and HeLa cell lysates and GST‐LC3C or GST‐GABARAP‐L2 immunoprecipitations.
- E
SDS–PAGE and Western blot of HEK293T cell lysates transfected with full‐length TBK1, a C‐terminal truncation mutant (TBK1 ΔC), a kinase‐dead version (TBK1 K38A), and GST‐LC3C or GST‐GABARAP‐L2 immunoprecipitations.
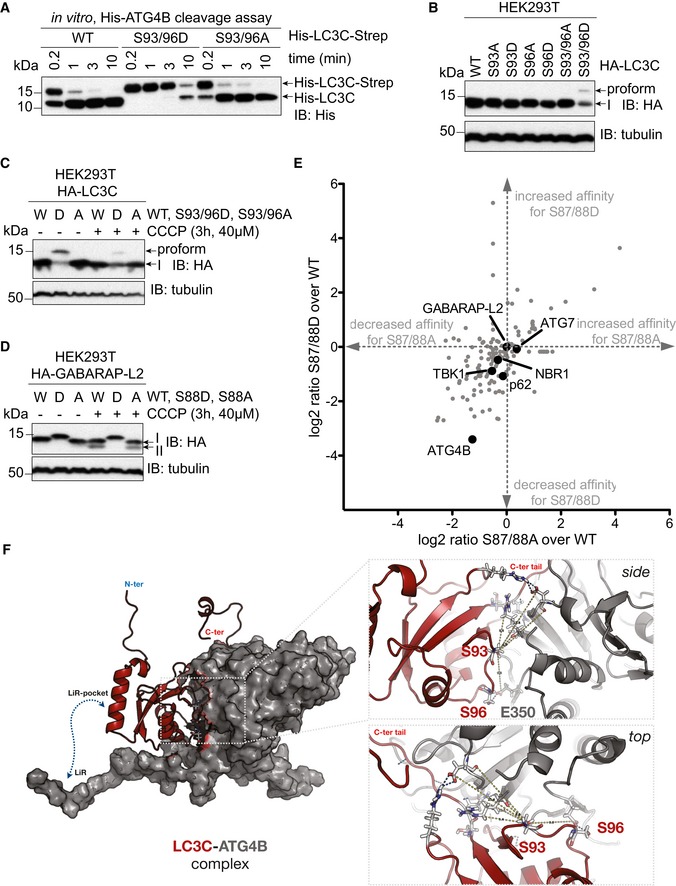
- A
SDS–PAGE and Western blot of in vitro ATG4 cleavage assay. Purified double‐tagged His‐LC3C‐Strep WT and mutants were incubated with ATG4B for indicated time points. LC3C S93/96D mutation slows down C‐terminal cleavage of LC3C by ATG4B.
- B
SDS–PAGE and Western blot of HEK293T cell lysates transfected with LC3C WT or mutants. S93/96D mutation of LC3C impedes cleavage of pro‐LC3C by endogenous ATG4s.
- C, D
SDS–PAGE and Western blot of HEK293T cell lysates transfected with LC3C WT or mutants (C) or GABARAP‐L2 WT or mutants (D). Cells were left untreated or treated with 40 μM CCCP for 3 h to induce mitophagy.
- E
Mass spectrometry interactome analysis of HEK293T cells with overexpressed HA‐GABARAP‐L2 WT, S87/88A, and S87/88D. Data were analyzed with Perseus, and binding partners that specifically bind GABARAP‐L2 in comparison with mock IP are depicted (n = 3 biological replicates).
- F
Full‐length LC3C (red cartoon) binds to ATG4B (gray surface) with its C‐terminal tail accessible to the active site of ATG4B. Phosphorylation of LC3C at S93 and S96 (sticks) affects binding to ATG4B. Zoom‐up showing the side and top view of LC3C‐ATG4B interface. S96 of LC3C and E350 of ATG4B form direct hydrogen bonds across the interface. S93 position is central to a network of polar interactions (blue dashed lines; side chains shown as sticks) across the interface.
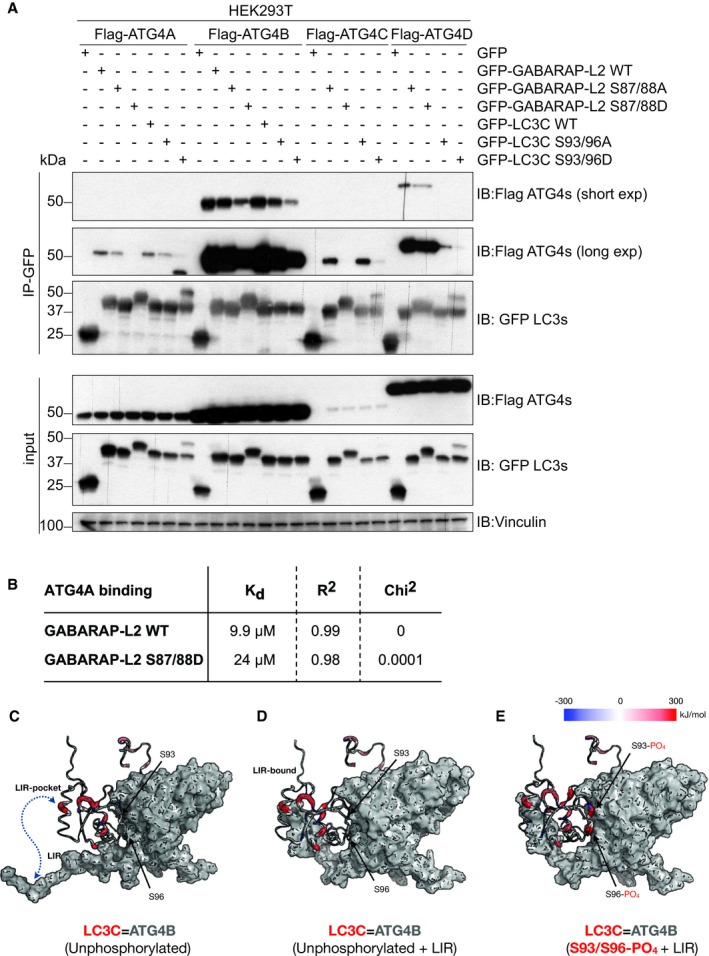
- A
SDS–PAGE and Western blot of HEK293T cell lysates and GFP immunoprecipitations. Cells were transfected with Flag‐tagged ATG4A, ATG4B, ATG4C, or ATG4D and GFP‐tagged LC3C or GABARAP‐L2 WT or mutants and lysates used for GFP IPs. S93/96D mutation of LC3C and S87/88D mutation of GABARAP‐L2 impede binding to ATG4A, ATG4B, ATG4C, and ATG4D.
- B
Bio‐Layer Interferometry measurement of GST‐ATG4A binding to His‐GABARAP‐L2 WT and S87/88D.
- C–E
(C) WT LC3C‐ATG4B complex (D) with additional LIR interactions and (E) with phosphorylated LC3C residues (S93 and S96) subjected to MD simulations and binding free energy computations using MM‐PBSA (see Materials and Methods) approach. A residue‐wise decomposition of the total binding free energy mapped onto the LC3C structure displays locally favorable (blue), neutral (white), and unfavorable (red) residue interaction with ATG4B (gray surface). S93 and S96 positions in WT complexes contribute favorably (blue), whereas in the phosphorylated complex they contribute unfavorably (red) toward complex formation. The thickness of the backbone scales linearly with the binding energy of LC3C‐ATG4B complexes.
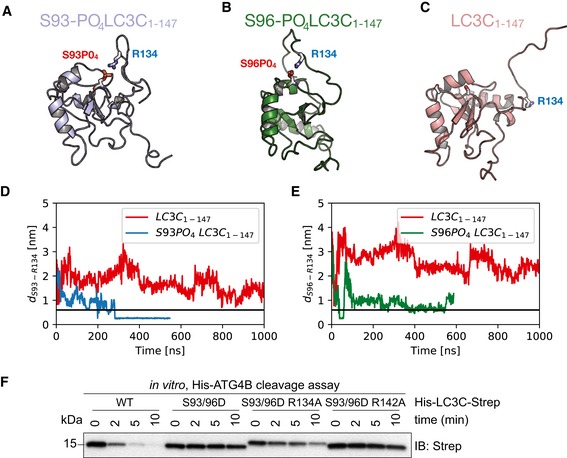
- A–C
Representative snapshots from all‐atom MD simulations (see [Link], [Link], [Link]) of unphosphorylated (red), S93‐PO4 LC3C (blue), and S96‐PO4 LC3C (green).
- D, E
Salt‐bridge formation dynamics in MD simulations represented by the time‐dependent minimum distance between side‐chain heavy atoms of R134 to (D) S93 and (E) S96 in phosphorylated (blue) and unphosphorylated (red) LC3C simulations. The horizontal black line (0.6 nm) indicates the cut‐off distance for stable electrostatic contact interactions.
- F
SDS–PAGE and Western blot of in vitro ATG4 cleavage assay. Purified double‐tagged His‐LC3C‐Strep WT and mutants were incubated with ATG4B in buffer for indicated time points. LC3C S93/96D R134A mutation enables C‐terminal cleavage of LC3C by ATG4B.
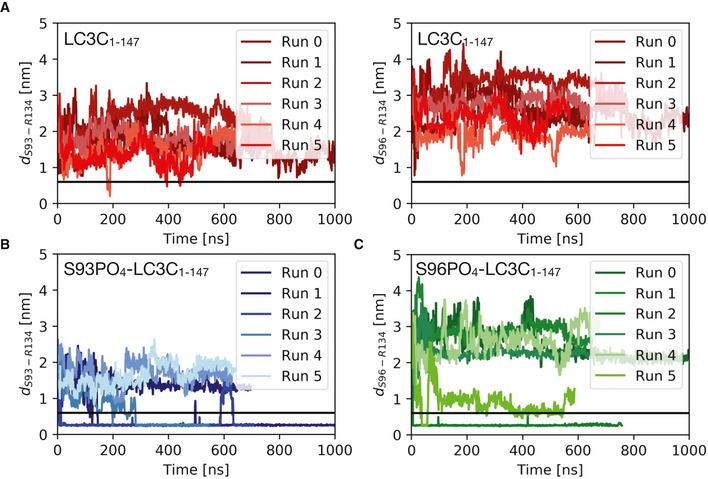
- A–C
Plots of the minimum distance between R134 and S93/S96 report on intramolecular salt‐bridge formation in MD simulations of the free LC3C with (A) unphosphorylated S93 and S96, (B) S93‐PO4, and (C) S96‐PO4. Replicas (n = 6) were initiated with different initial velocities after equilibration (at 310 K, 1 bar, and 150 mM NaCl) to capture variations in the C‐terminal tail dynamics in phosphorylated and unphosphorylated LC3C. They consistently show the formation of intramolecular salt bridges between the phosphorylated serine residues and the positively charged R134 in the C‐terminal tail. The horizontal black line (0.6 nm) indicates the cut‐off distance for stable electrostatic contact interactions.
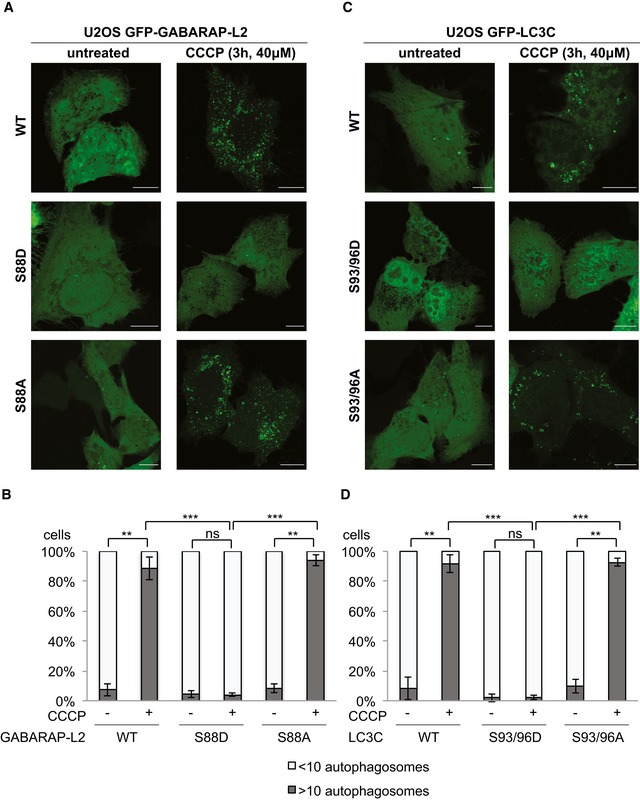
- A–D
U2OS cells were transfected with GFP‐GABARAP‐L2 (A) or GFP‐LC3C (C) WT or mutants and HA‐Parkin. Mitophagy was induced by the addition of 40 μM CCCP for 3 h. WT, S87/88A GABARAP‐L2 and WT, S93/96A LC3C localize to autophagosomes, whereas S87/88D GABARAP‐L2 and S93/96D LC3C remain dispersed in the cytosol. Scale bar represents 10 μm. (B, D) GFP‐expressing cells were counted and segregated into classes with greater and less than 10 autophagosomes per cell. Data are presented as mean ± SD, n = 3 biological replicates, **P < 0.01, ***P < 0.001, ns = not significant, as analyzed by Student's t‐test.
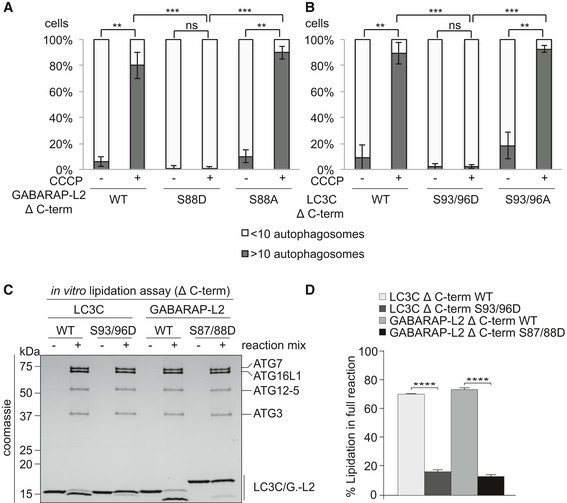
- A, B
U2OS cells were transfected with GFP‐GABARAP‐L2 Δ C‐terminal (A) or GFP‐LC3C Δ C‐terminal (B) WT or mutants and HA‐Parkin. Mitophagy was induced by the addition of 40 μM CCCP for 3 h. GFP‐expressing cells were counted and segregated into classes with greater and less than 10 autophagosomes per cell. Data are presented as mean ± SD, n = 3 biological replicates, **P < 0.01, ***P < 0.001 as analyzed by Student's t‐test.
- C
In vitro lipidation reactions containing 10 μM LC3C WT, LC3C S93/96D, GABARAP‐L2 WT, or GABARAP‐L2 S87/88D incubated with or without reaction mix (0.5 μM hATG7, 1 μM hATG3, 0.25 μM hATG12‐ATG5‐ATG16L1, 3 mM lipid (sonicated liposomes composed of 10 mol% bl‐PI, 55 mol% DOPE, and 35 mol% POPC), 1 mM DTT, and 1 mM ATP) were incubated at 30°C for 90 min. The reactions were analyzed by SDS–PAGE and visualized by Coomassie blue stain.
- D
The extent of lipidation in (C) was quantified and plotted as percentage of total protein (conjugated and unconjugated). Data are presented as mean ± SEM, n = 3 biological replicates, ****P < 0.0001, as analyzed by one‐way ANOVA followed by Bonferroni's multiple comparison test.
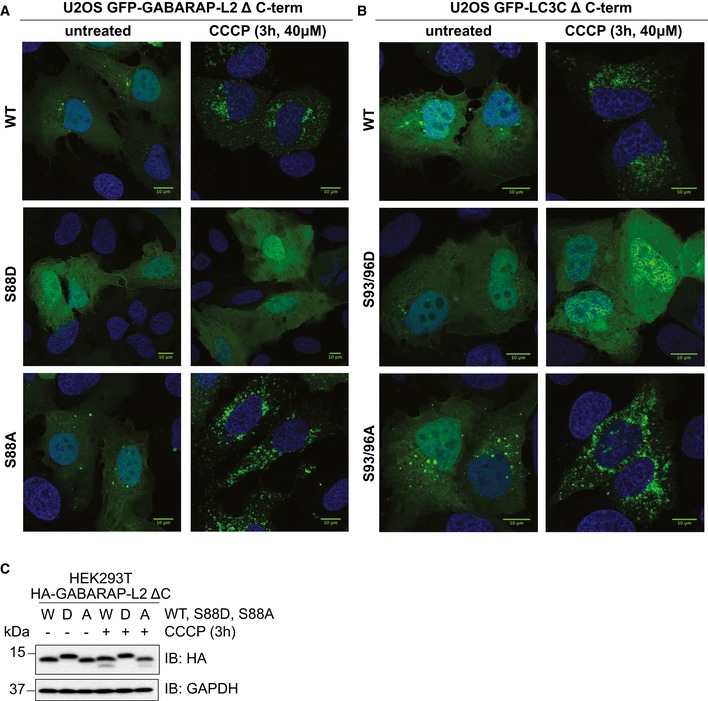
- A, B
U2OS cells were transfected with GFP‐GABARAP‐L2 Δ C‐terminal (A) or GFP‐LC3C Δ C‐terminal (B) WT or mutants and HA‐Parkin. Mitophagy was induced by the addition of 40 μM CCCP for 3 h. WT, S87/88A Δ C‐terminal GABARAP‐L2 and WT, S93/96A Δ C‐terminal LC3C localize to autophagosomes, whereas S87/88D Δ C‐terminal GABARAP‐L2 and S93/96D Δ C‐terminal LC3C remain dispersed in the cytosol. Scale bar represents 10 μm.
- C
SDS–PAGE and Western blot of HEK293T cell lysates transfected with HA‐GABARAP‐L2 Δ C‐terminal WT or mutants. Cells were left untreated or treated with 40 μM CCCP for 3 h to induce mitophagy.
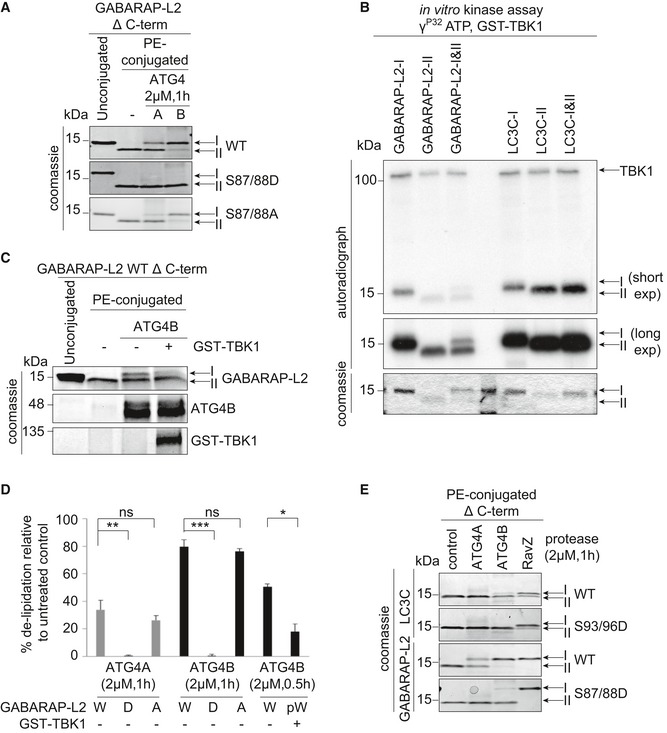
GABARAP‐L2 WT‐, S87/88D‐, or S87/88A‐conjugated liposomes were treated or not with 2 μM ATG4A or ATG4B for 1 h at 37°C. Samples were then subjected to SDS–PAGE together with unconjugated GABARAP‐L2‐I.
Coomassie stain and autoradiography of SDS–PAGE after an in vitro kinase assay with GST‐TBK1, PE‐conjugated (II), and unconjugated (I) GABARAP‐L2 and LC3C.
PE‐conjugated GABARAP‐L2‐II WT was incubated in phosphorylation assay buffer in the presence or absence of GST‐TBK1 for 4 h and treated or not with 2 μM ATG4B for 30 min. Samples were then subjected to SDS–PAGE together with unconjugated GABARAP‐L2‐I.
The extent of de‐lipidation in (A) and (C) was quantified and plotted as percentage of total protein (conjugated and unconjugated). W = WT, D = S87/88D, A = S87/88A, pW = TBK1‐phosphorylated WT GABARAP‐L2. Data are presented as mean ± SD, n = 3 biological replicates, *P < 0.05, **P < 0.01, ***P < 0.001, ns = not significant, as analyzed by Student's t‐test.
LC3C WT‐, LC3C S93/96D‐, GABARAP‐L2 WT‐, or GABARAP‐L2 S87/88D‐conjugated liposomes were treated or not with 2 μM ATG4A, ATG4B, or RavZ for 1 h at 37°C. Samples were then subjected to SDS–PAGE.
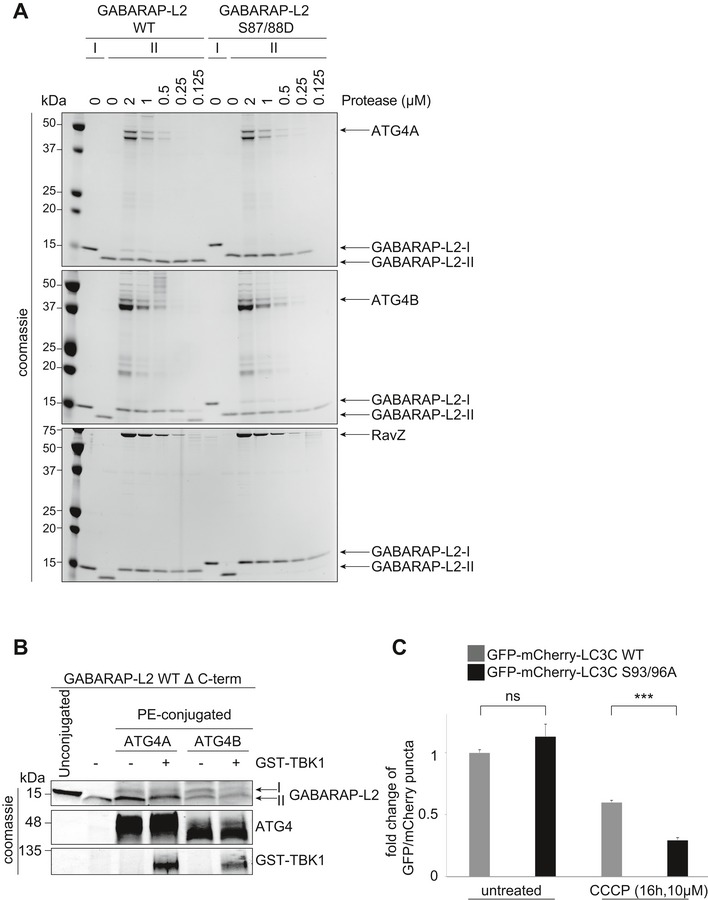
GABARAP‐L2 WT‐ or S87/88D‐conjugated liposomes (II) were treated or not with different amounts (2–0.125 μM) of ATG4A, ATG4B, or RavZ for 1 h at 37°C. Samples were then subjected to SDS–PAGE together with unconjugated GABARAP‐L2 (I).
PE‐conjugated GABARAP‐L2‐II WT was incubated in phosphorylation assay buffer in the presence or absence of GST‐TBK1 for 4 h and treated or not with 2 μM ATG4A or ATG4B for 30 min. Samples were then subjected to SDS–PAGE together with unconjugated GABARAP‐L2‐I.
HeLa cells expressing doxycycline‐inducible Parkin and stable mCherry‐GFP‐LC3C WT or S93/96A were treated with 1 μg/ml doxycycline and 10 μM CCCP for 16 h. mCherry and GFP‐positive puncta were assessed by microscopy (Yokogawa CQ1), and the ratio of GFP over mCherry puncta per cell was calculated for each sample. Data are presented as mean ± SD, > 500 cells/condition, n = 3 biological replicates.
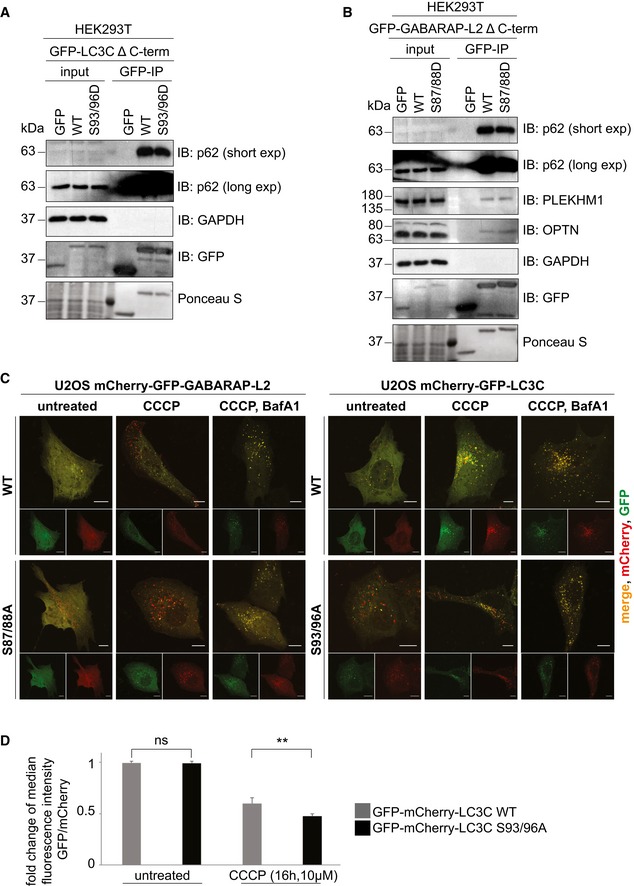
- A, B
SDS–PAGE and Western blot of HEK293T cell lysates and GFP IPs. (A) Cells were transfected with GFP, GFP‐LC3C Δ C‐terminal WT or S93/96D, and lysates used for GFP IPs. WT and S93/96D LC3C bind endogenous p62. (B) Cells were transfected with GFP, GFP‐GABARAP‐L2 Δ C‐terminal WT or S87/88D, and lysates used for GFP IPs. WT and S87/88D GABARAP‐L2 bind endogenous p62, optineurin (OPTN), and PLEKHM1.
- C
U2OS cells were seeded onto coverslips and transfected with mCherry‐GFP‐LC3C WT or S93/96A or mCherry‐GFP‐GABARAP‐L2 WT or S87/88A. Mitophagy was induced by the addition of 40 μM CCCP for 3 h, and lysosomal degradation of GFP was blocked by the addition of 200 nM bafilomycin A1 for 3 h. Scale bar represents 10 μm.
- D
HeLa cells expressing doxycycline‐inducible Parkin and stable mCherry‐GFP‐LC3C WT or S93/96A were treated with 1 μg/ml doxycycline and 10 μM CCCP for 16 h. mCherry and GFP fluorescence was measured by FACS, and the fold change of median fluorescence intensity of GFP over mCherry was calculated for each sample. Data are presented as mean ± SD. n = 6 biological replicates, **P < 0.01, ns = not significant, as analyzed by Student's t‐test.
References
-
- Dikic I (2017) Proteasomal and autophagic degradation systems. Annu Rev Biochem 86: 193–224 - PubMed
-
- Nakatogawa H (2013) Two ubiquitin‐like conjugation systems that mediate membrane formation during autophagy. Essays Biochem 55: 39–50 - PubMed
-
- Stolz A, Ernst A, Dikic I (2014) Cargo recognition and trafficking in selective autophagy. Nat Cell Biol 16: 495–501 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
