

Histamine H1 receptor on astrocytes and neurons controls distinct aspects of mouse behaviour
- PMID: 31712580
- PMCID: PMC6848115
- DOI: 10.1038/s41598-019-52623-6
Histamine H1 receptor on astrocytes and neurons controls distinct aspects of mouse behaviour
Abstract
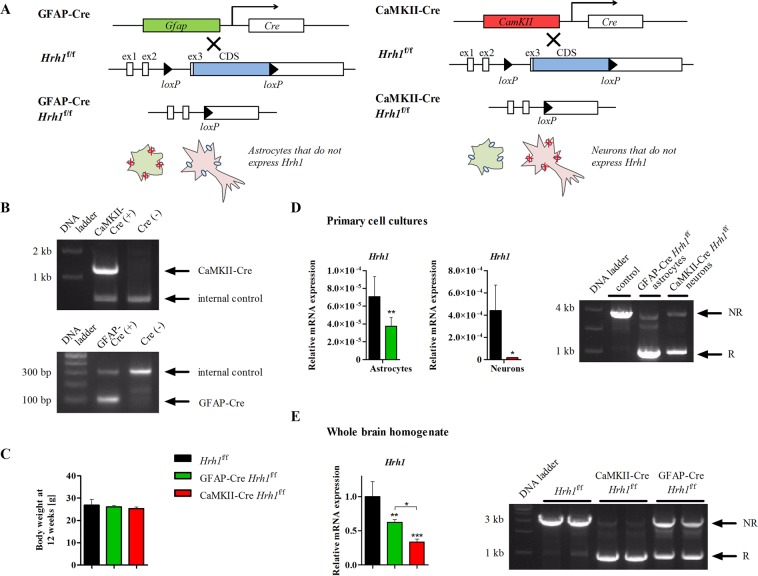
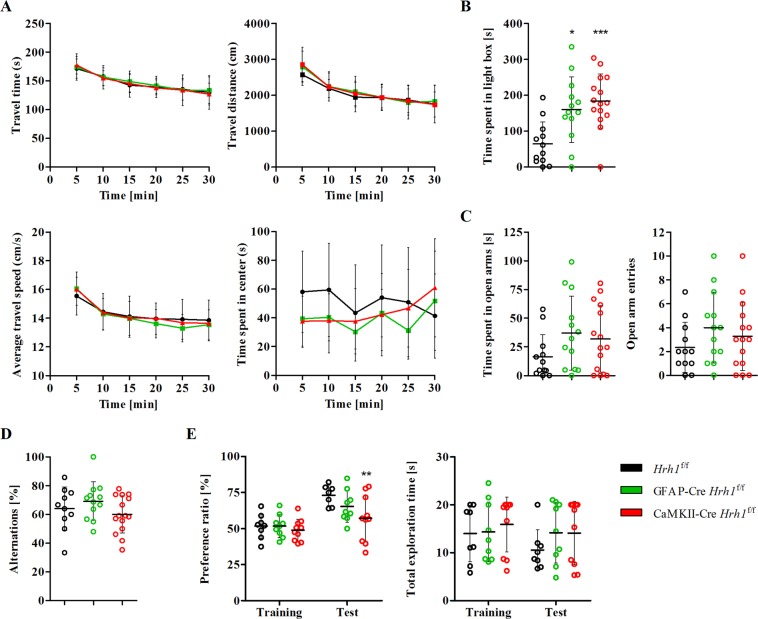
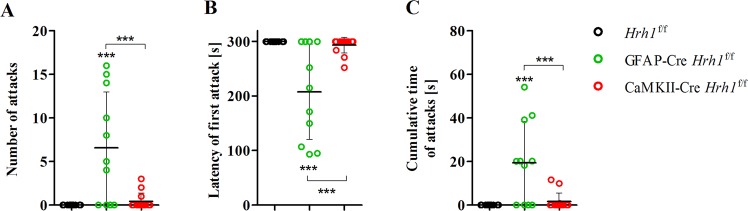
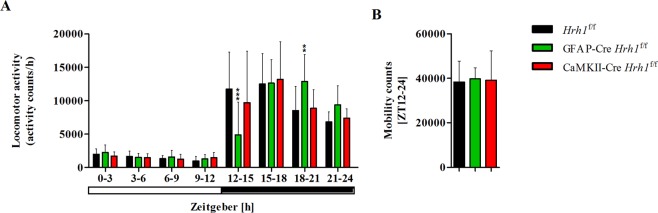
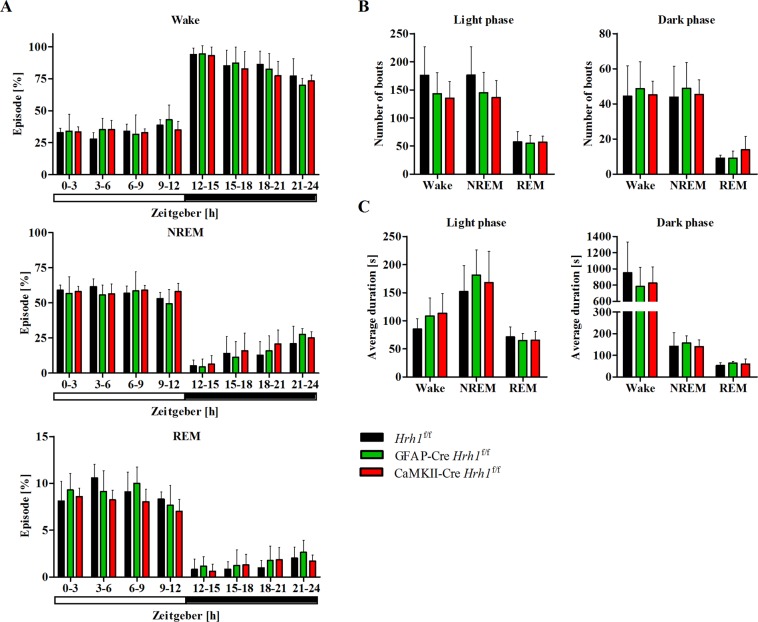
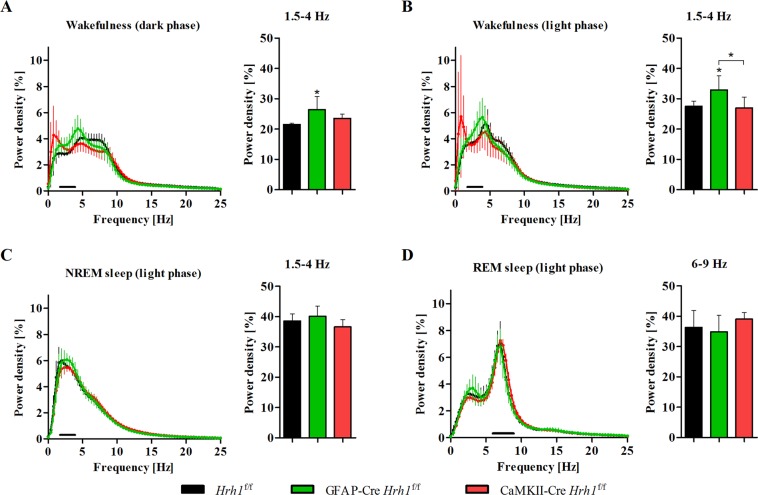
Histamine is an important neurotransmitter that contributes to various processes, including the sleep-wake cycle, learning, memory, and stress responses. Its actions are mediated through histamine H1-H4 receptors. Gene knockout and pharmacological studies have revealed the importance of H1 receptors in learning and memory, regulation of aggression, and wakefulness. H1 receptors are abundantly expressed on neurons and astrocytes. However, to date, studies selectively investigating the roles of neuronal and astrocytic H1 receptors in behaviour are lacking. We generated novel astrocyte- and neuron-specific conditional knockout (cKO) mice to address this gap in knowledge. cKO mice showed cell-specific reduction of H1 receptor gene expression. Behavioural assessment revealed significant changes and highlighted the importance of H1 receptors on both astrocytes and neurons. H1 receptors on both cell types played a significant role in anxiety. Astrocytic H1 receptors were involved in regulating aggressive behaviour, circadian rhythms, and quality of wakefulness, but not sleep behaviour. Our results emphasise the roles of neuronal H1 receptors in recognition memory. In conclusion, this study highlights the novel roles of H1 receptors on astrocytes and neurons in various brain functions.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
