

Calcium ions in the aquatic environment drive planarians to food
- PMID: 31720007
- PMCID: PMC6836377
- DOI: 10.1186/s40851-019-0147-x
Calcium ions in the aquatic environment drive planarians to food
Abstract
Background: Even subtle changes in environmental factors can exert behavioral effects on creatures, which may alter interspecific interactions and eventually affect the ecosystem. However, how changes in environmental factors impact complex behaviors regulated by neural processes is largely unknown. The freshwater planarian Dugesia japonica, a free-living flatworm, displays distinct behavioral traits mediated by sensitive perception of environmental cues. Planarians are thus useful organisms for examining interactions between environmental changes and specific behaviors of animals.
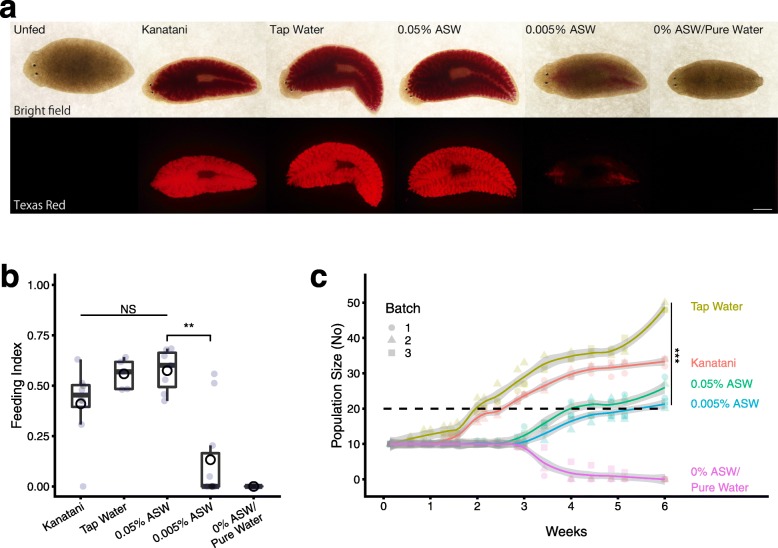
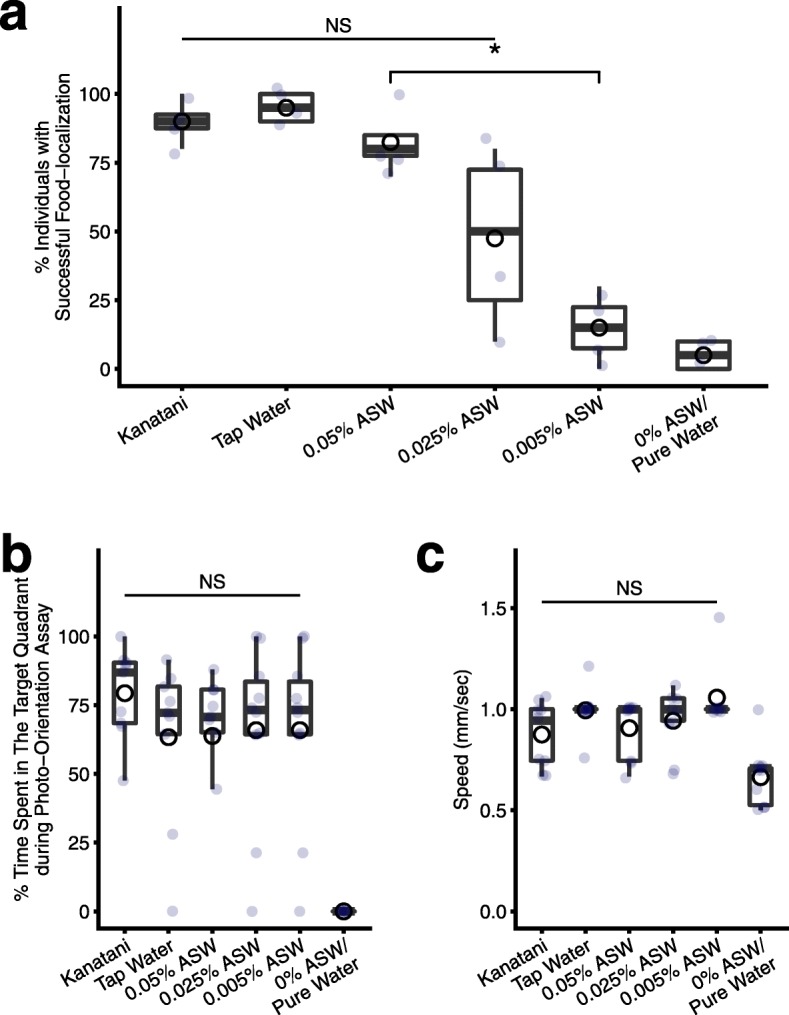
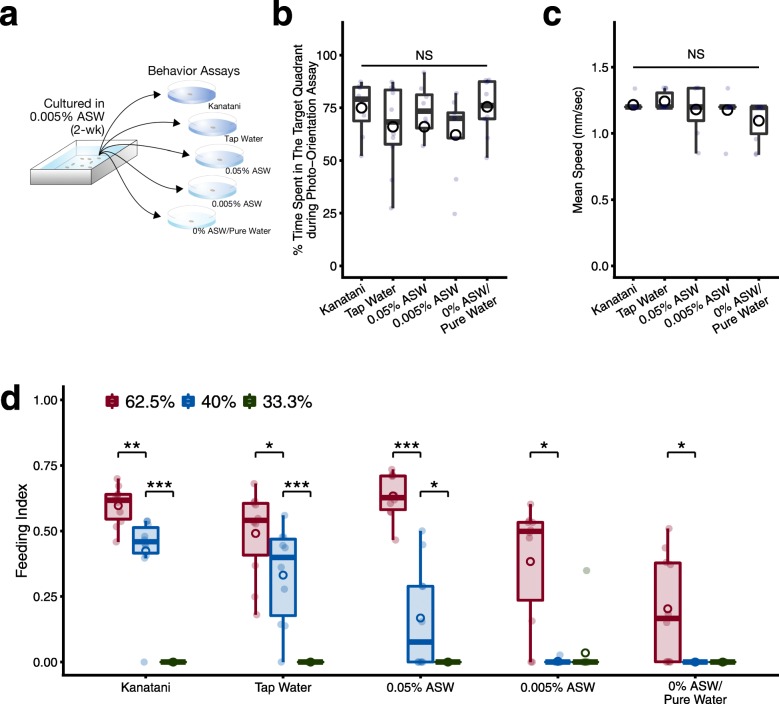
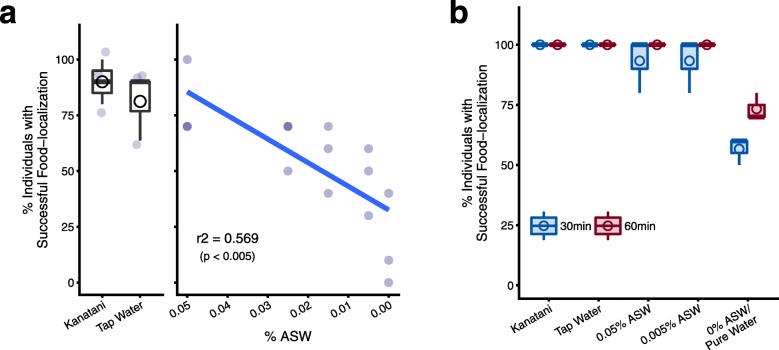
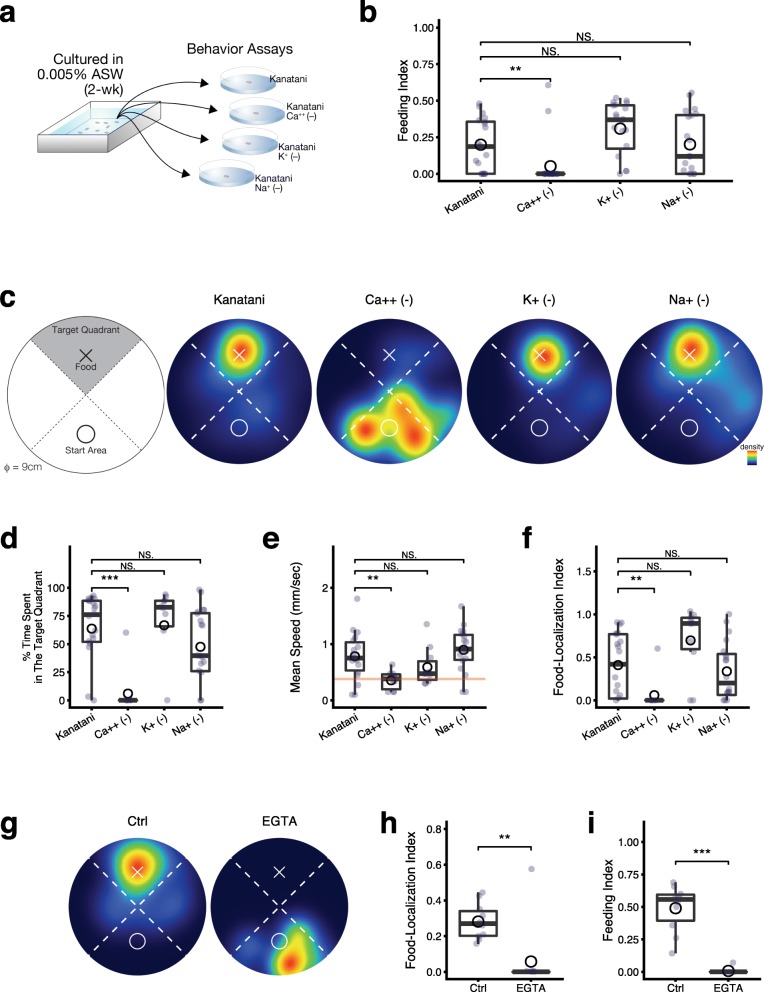
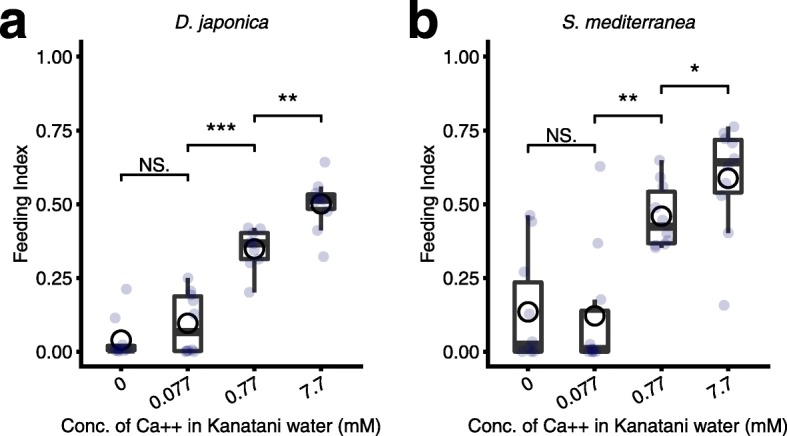
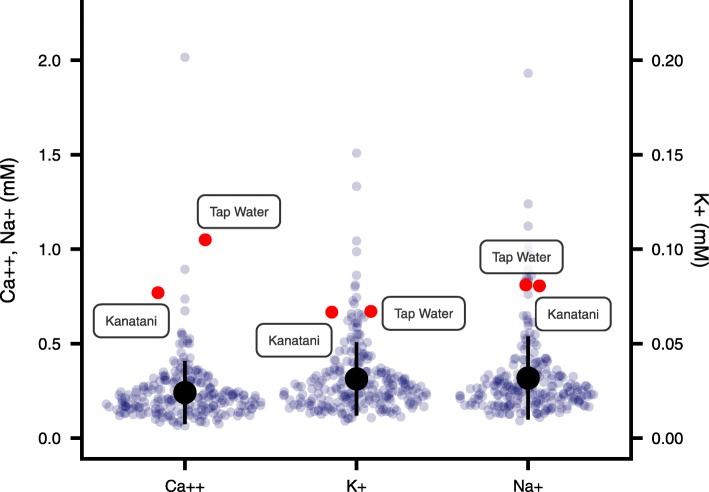
Results: Here we found that feeding behavior was suppressed when the concentration of ions in the breeding water was low, while other behaviors were unaffected, resulting in differences in population size. Notably, the decline in feeding behavior was reversed in an ion-concentration-dependent manner soon after the planarians were moved to ion-containing water, which suggests that ions in environmental water rapidly promote feeding behavior in planarians. Moreover, the concentration of ions in the environmental water affected the feeding behavior by modulating the sensitivity of the response to foods. Finally, we found that calcium ions in the aquatic environment were required for the feeding behavior, and exposure to higher levels of calcium ions enhanced the feeding behavior, showing that there was a good correlation between the concentration of calcium ions and the responsiveness of planarians to foods.
Conclusions: Environmental calcium ions are indispensable for and potentiate the activity level of the feeding behavior of planarians. Our findings suggest that the ions in the aquatic environment profoundly impact the growth and survival of aquatic animals via modulating their neural activities and behaviors.
Keywords: Environment-responsive behavior; Environmental changes; Feeding behavior; Planarian; Quantitative behavioristics; Responsive sensitivity.
© The Author(s). 2019.
Conflict of interest statement
Competing interestsThe authors declare no competing financial or non-financial Interests.
Figures
Similar articles
-
The pharyngeal nervous system orchestrates feeding behavior in planarians.Sci Adv. 2020 Apr 8;6(15):eaaz0882. doi: 10.1126/sciadv.aaz0882. eCollection 2020 Apr. Sci Adv. 2020. PMID: 32285000 Free PMC article.
-
Toxic effects of sodium dodecyl sulfate on planarian Dugesia japonica.PeerJ. 2023 Jul 10;11:e15660. doi: 10.7717/peerj.15660. eCollection 2023. PeerJ. 2023. PMID: 37456884 Free PMC article.
-
Antioxidant responses and lipid peroxidation can be used as sensitive indicators for the heavy metals risk assessment of the Wei River: a case study of planarian Dugesia Japonica.Biomarkers. 2021 Feb;26(1):55-64. doi: 10.1080/1354750X.2020.1854347. Epub 2020 Dec 21. Biomarkers. 2021. PMID: 33225756
-
Regeneration in an evolutionarily primitive brain--the planarian Dugesia japonica model.Eur J Neurosci. 2011 Sep;34(6):863-9. doi: 10.1111/j.1460-9568.2011.07819.x. Eur J Neurosci. 2011. PMID: 21929621 Review.
-
Neuropharmacology and behavior in planarians: translations to mammals.Comp Biochem Physiol C Toxicol Pharmacol. 2008 May;147(4):399-408. doi: 10.1016/j.cbpc.2008.01.009. Epub 2008 Feb 1. Comp Biochem Physiol C Toxicol Pharmacol. 2008. PMID: 18294919 Review.
Cited by
-
Establishment of a new method to isolate viable x-ray-sensitive cells from planarian by fluorescence-activated cell sorting.Dev Growth Differ. 2023 Dec;65(9):577-590. doi: 10.1111/dgd.12886. Epub 2023 Sep 5. Dev Growth Differ. 2023. PMID: 37596847 Free PMC article.
-
Regeneration of Planarian Auricles and Reestablishment of Chemotactic Ability.Front Cell Dev Biol. 2021 Nov 26;9:777951. doi: 10.3389/fcell.2021.777951. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34901022 Free PMC article.
-
Isolation of planarian viable cells using fluorescence-activated cell sorting for advancing single-cell transcriptome analysis.Genes Cells. 2023 Nov;28(11):800-810. doi: 10.1111/gtc.13068. Epub 2023 Sep 18. Genes Cells. 2023. PMID: 37723830 Free PMC article.
-
The pharyngeal nervous system orchestrates feeding behavior in planarians.Sci Adv. 2020 Apr 8;6(15):eaaz0882. doi: 10.1126/sciadv.aaz0882. eCollection 2020 Apr. Sci Adv. 2020. PMID: 32285000 Free PMC article.
-
Spatial localization ability of planarians identified through a light maze paradigm.PLoS One. 2023 Jul 19;18(7):e0288118. doi: 10.1371/journal.pone.0288118. eCollection 2023. PLoS One. 2023. PMID: 37467232 Free PMC article.
References
-
- Kawakatsu M, Oki I, Tamura S. Taxonomy and geographical distribution of Dugesia japonica and D. ryukyuensis in the far east. Hydrobiologia. 1995;305:55–61. doi: 10.1007/BF00036363. - DOI
-
- New TR. An introduction to invertebrate conservation biology. Oxford: Oxford University Press; 1995.
-
- Reddien PW, Sanchez Alvarad A. Fundamentals of planarian regeneration. Annu Rev Cell Dev Biol. 2004;20:725–57. - PubMed
-
- Shibata N, Rouhana L, Agata K. Cellular and molecular dissection of pluripotent adult somatic stem cells in planarians. Dev Growth Differ. 2010;52:27–41. - PubMed
LinkOut - more resources
Full Text Sources
