

The histone variant H2A.Z in yeast is almost exclusively incorporated into the +1 nucleosome in the direction of transcription
- PMID: 31722407
- PMCID: PMC7145542
- DOI: 10.1093/nar/gkz1075
The histone variant H2A.Z in yeast is almost exclusively incorporated into the +1 nucleosome in the direction of transcription
Abstract
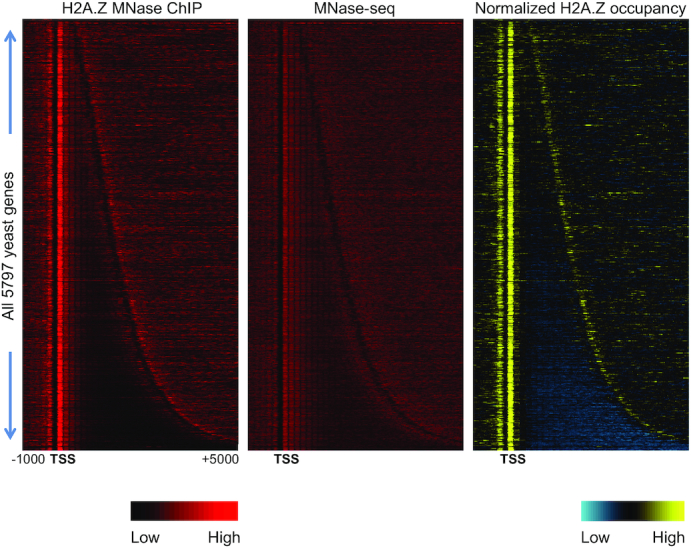
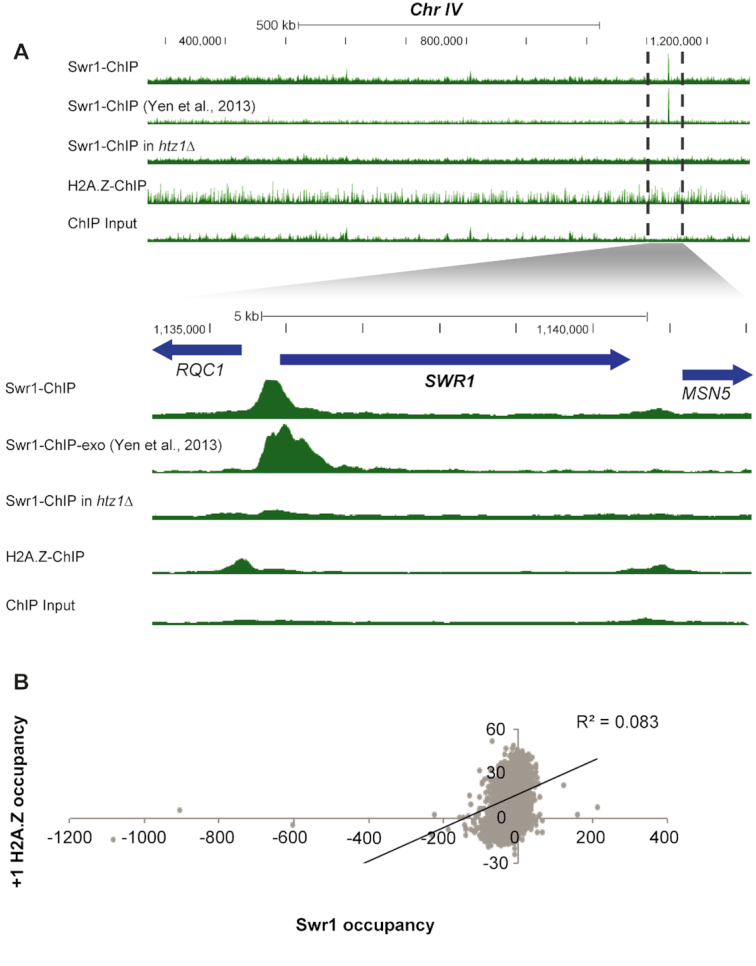
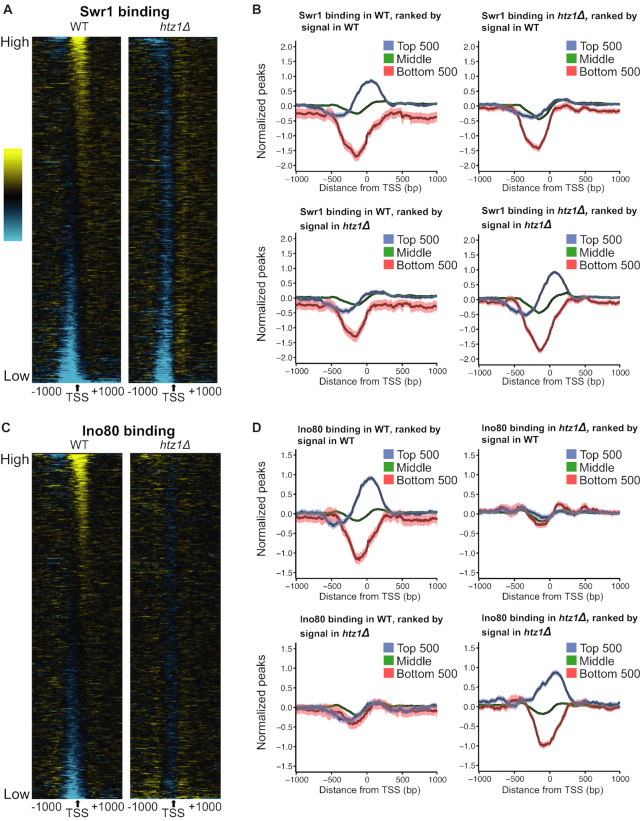
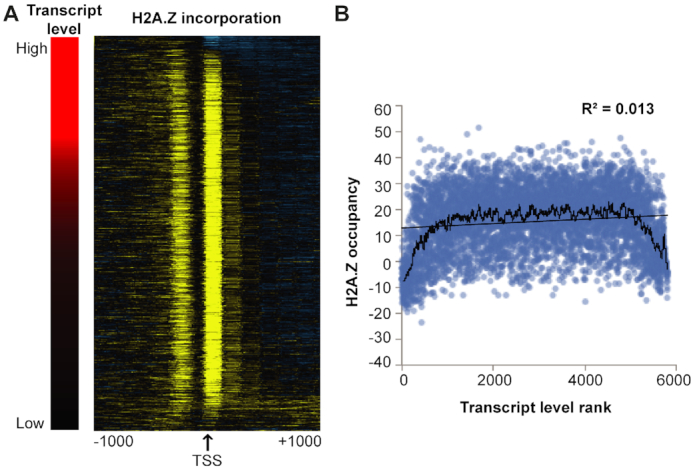
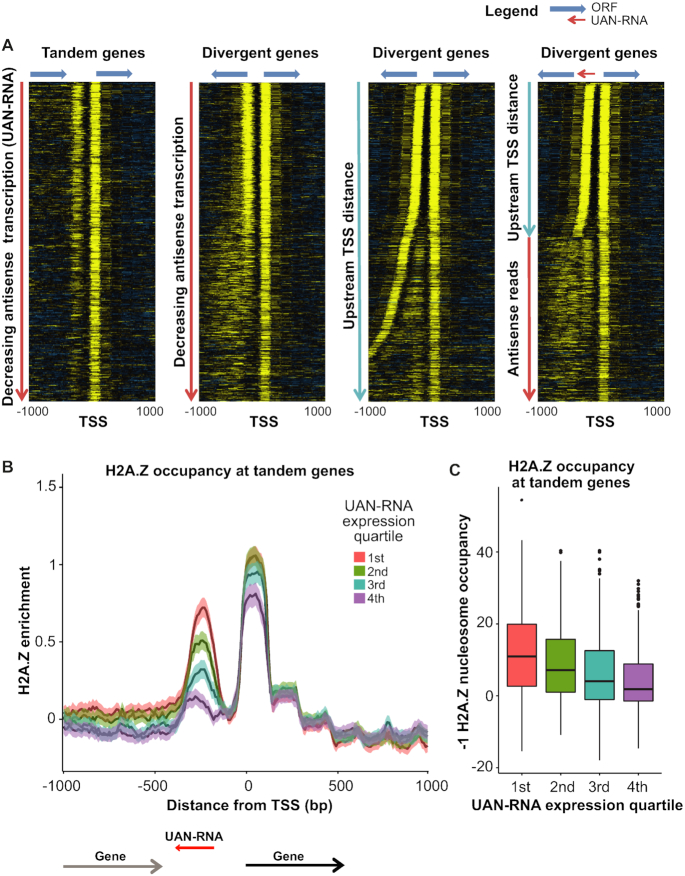
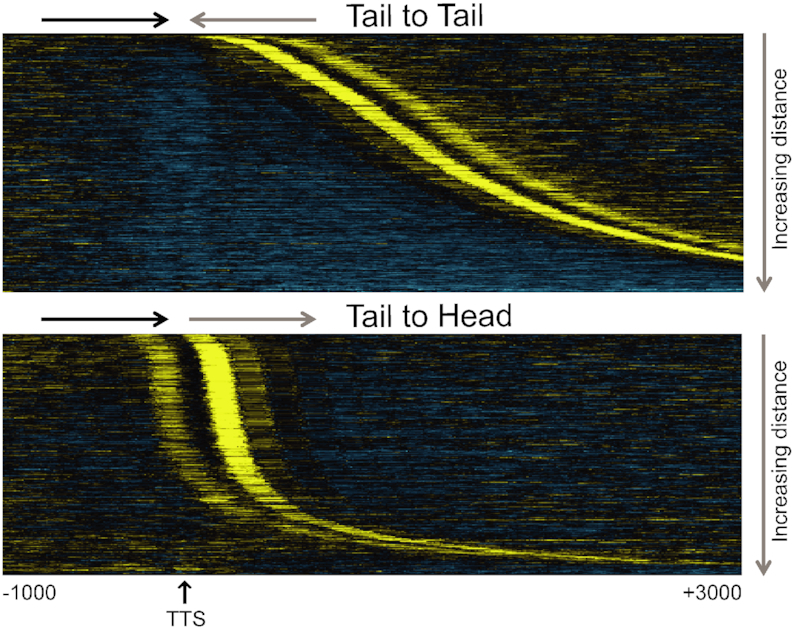
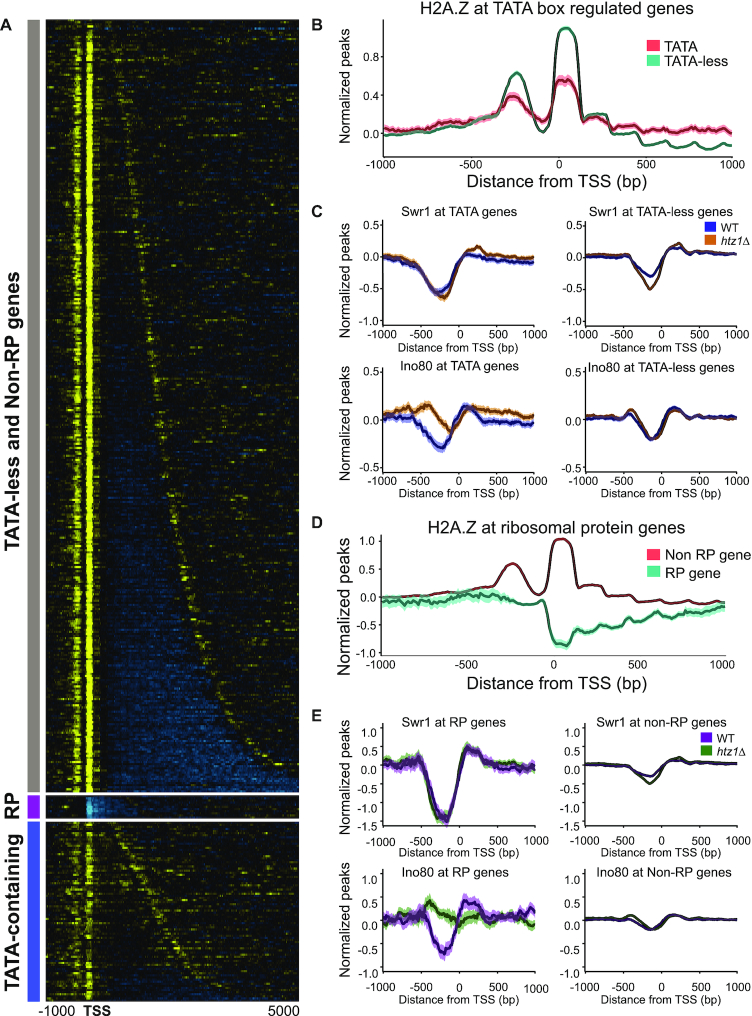
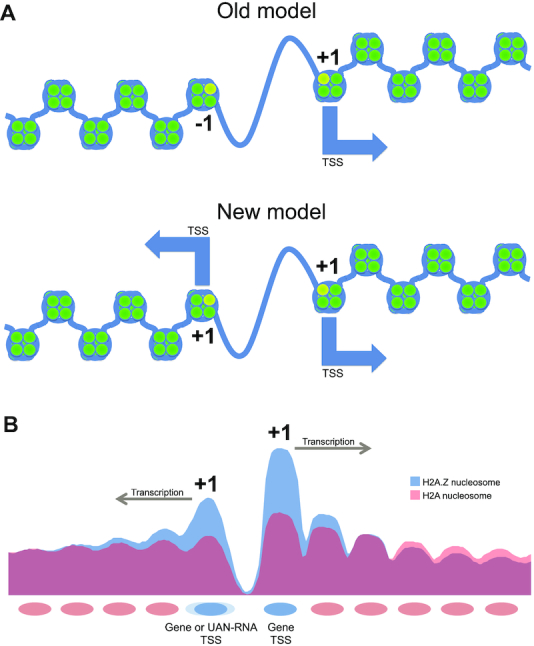
Transcription start sites (TSS) in eukaryotes are characterized by a nucleosome-depleted region (NDR), which appears to be flanked upstream and downstream by strongly positioned nucleosomes incorporating the histone variant H2A.Z. H2A.Z associates with both active and repressed TSS and is important for priming genes for rapid transcriptional activation. However, the determinants of H2A.Z occupancy at specific nucleosomes and its relationship to transcription initiation remain unclear. To further elucidate the specificity of H2A.Z, we determined its genomic localization at single nucleosome resolution, as well as the localization of its chromatin remodelers Swr1 and Ino80. By analyzing H2A.Z occupancy in conjunction with RNA expression data that captures promoter-derived antisense initiation, we find that H2A.Z's bimodal incorporation on either side of the NDR is not a general feature of TSS, but is specifically a marker for bidirectional transcription, such that the upstream flanking -1 H2A.Z-containing nucleosome is more appropriately considered as a +1 H2A.Z nucleosome for antisense transcription. The localization of H2A.Z almost exclusively at the +1 nucleosome suggests that a transcription-initiation dependent process could contribute to its specific incorporation.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Yuan G.C., Liu Y.J., Dion M.F., Slack M.D., Wu L.F., Altschuler S.J., Rando O.J.. Genome-scale identification of nucleosome positions in S. cerevisiae. Science. 2005; 309:626–630. - PubMed
-
- Albert I., Mavrich T.N., Tomsho L.P., Qi J., Zanton S.J., Schuster S.C., Pugh B.F.. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature. 2007; 446:572–576. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
