

Citrus PH4-Noemi regulatory complex is involved in proanthocyanidin biosynthesis via a positive feedback loop
- PMID: 31728522
- PMCID: PMC7031078
- DOI: 10.1093/jxb/erz506
Citrus PH4-Noemi regulatory complex is involved in proanthocyanidin biosynthesis via a positive feedback loop
Abstract
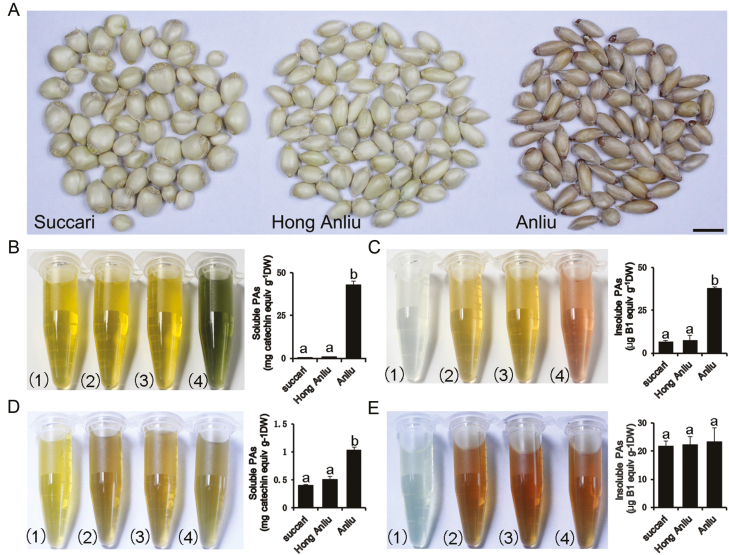
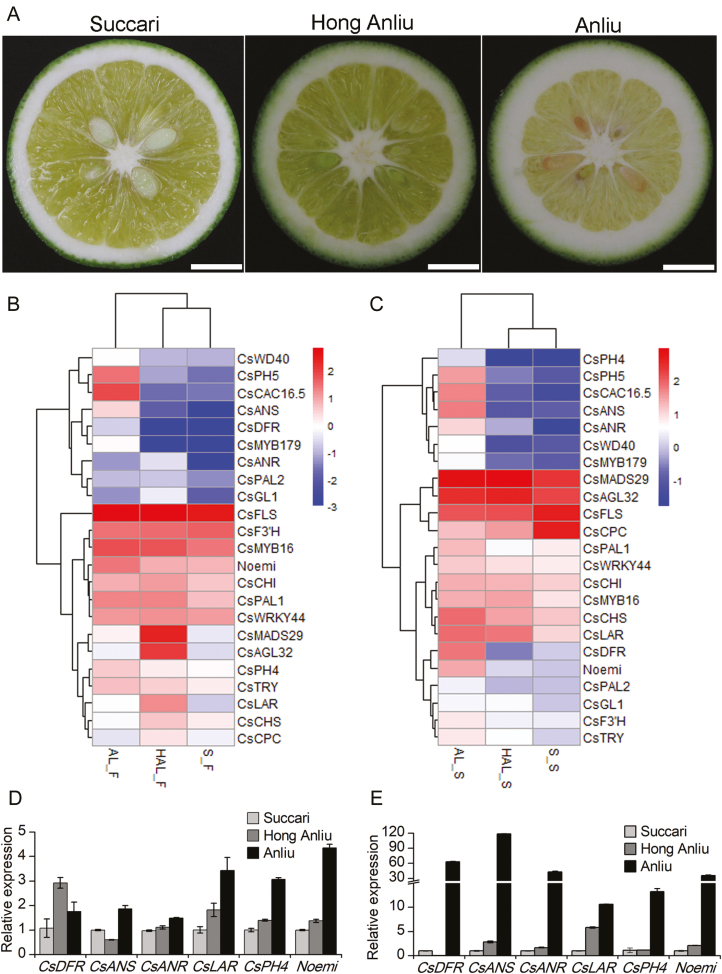
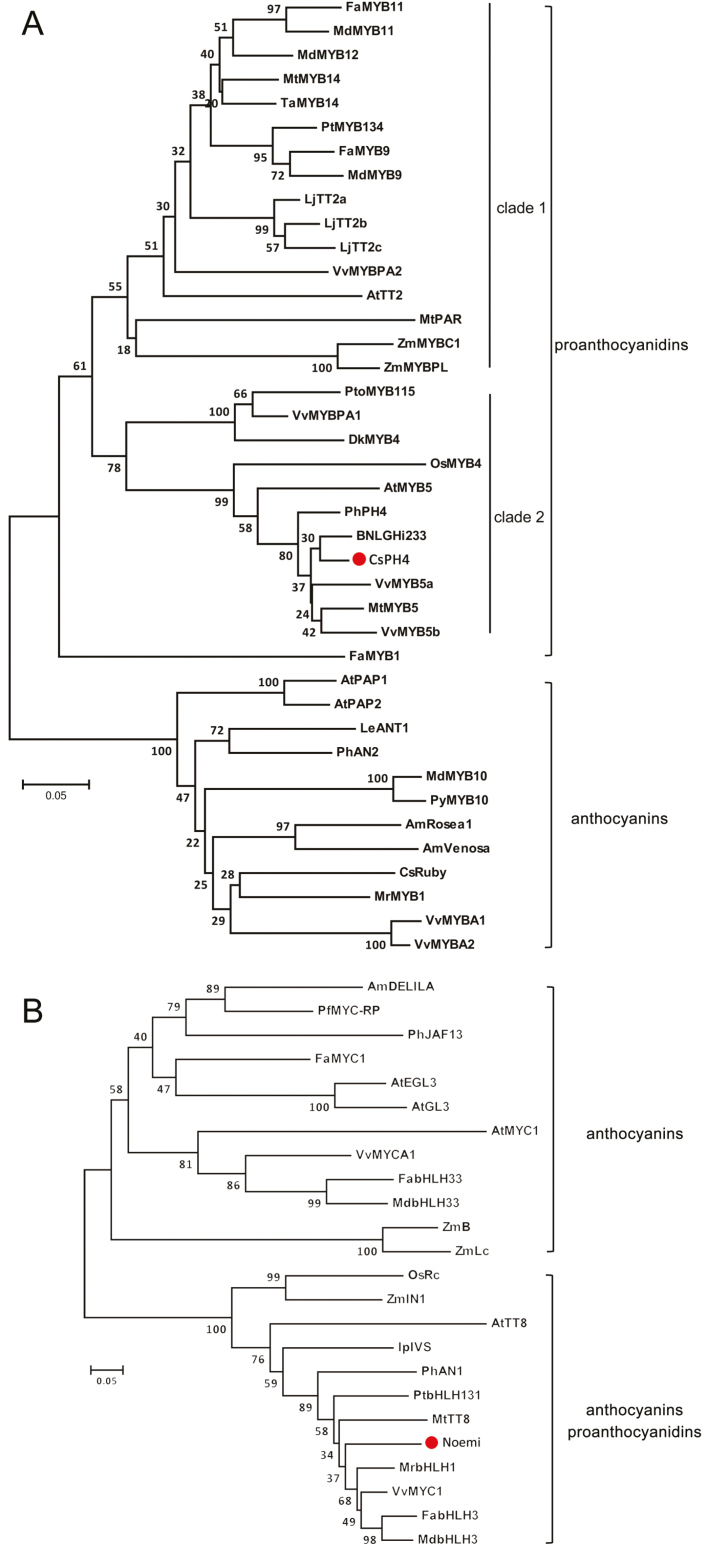
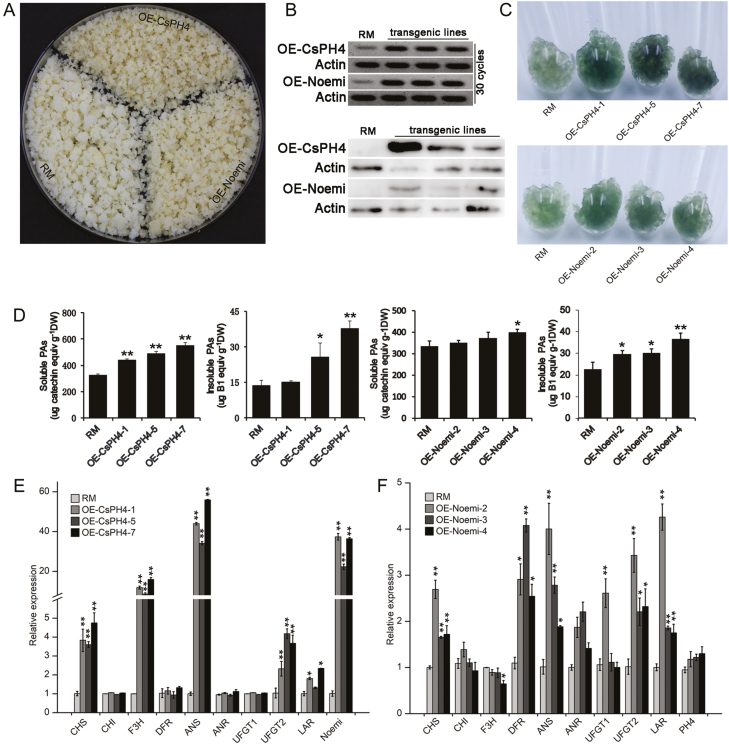
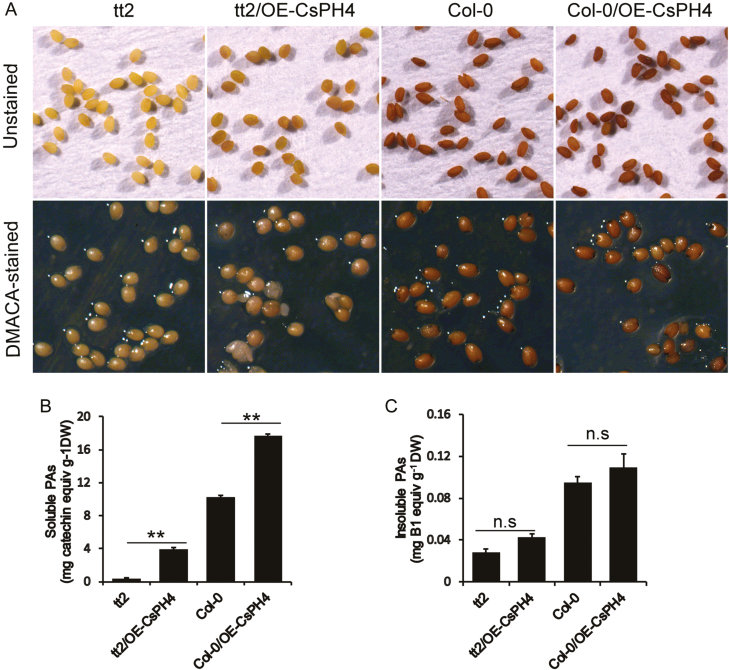
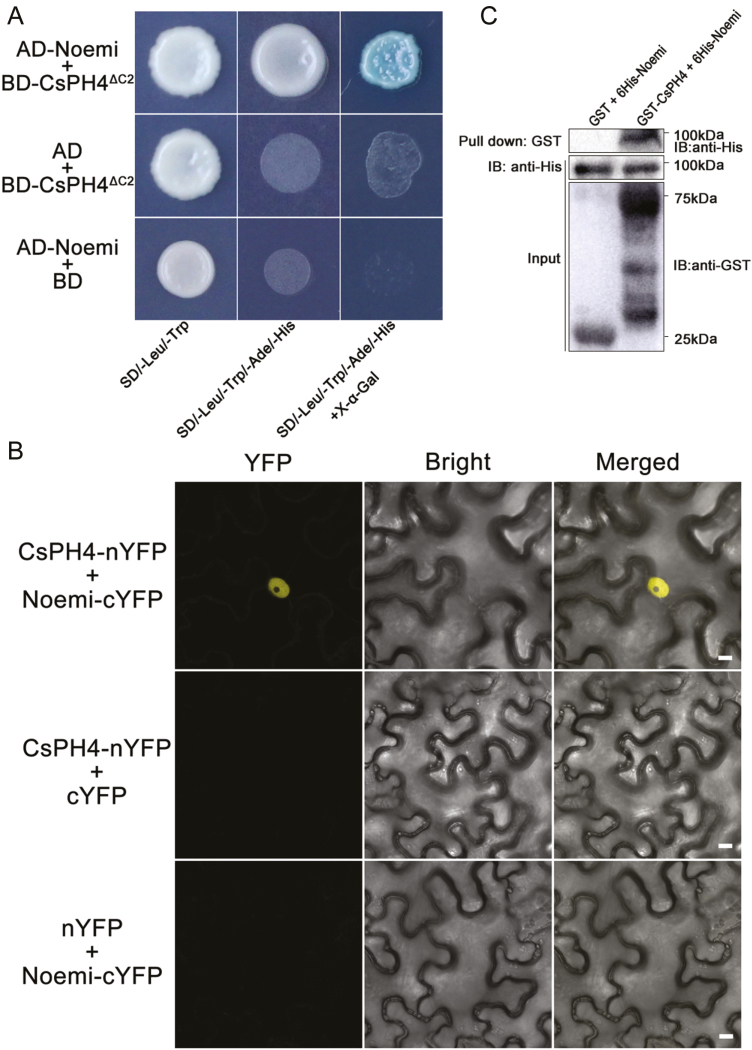
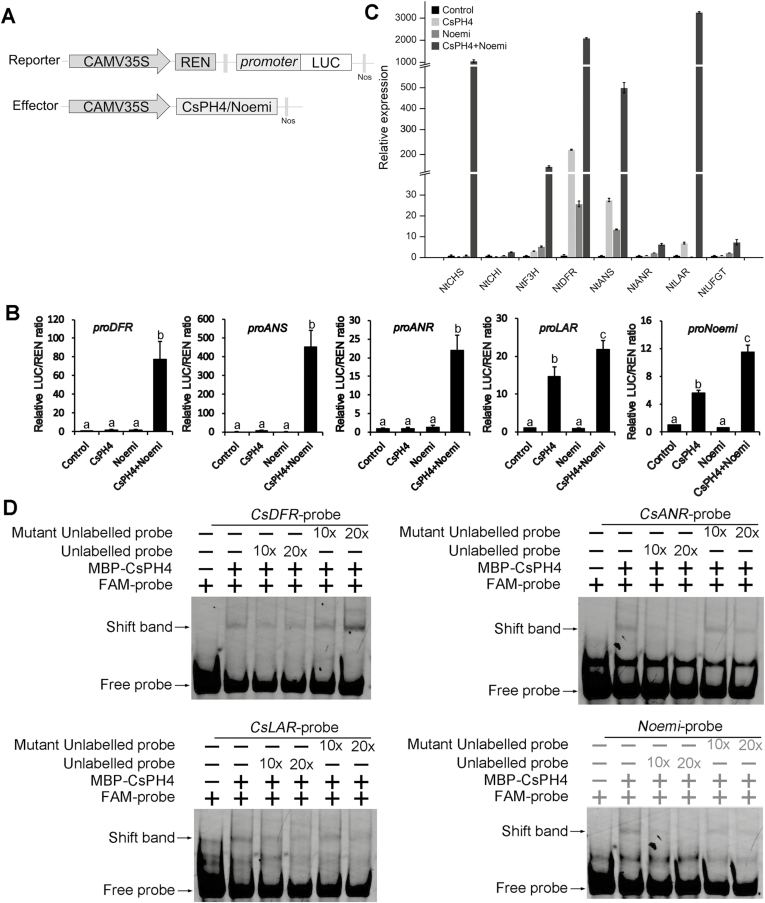
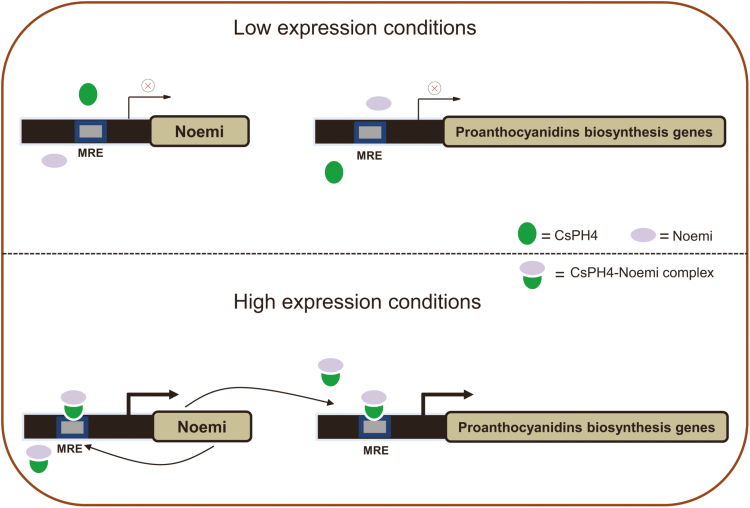
Proanthocyanidins (PAs; or condensed tannins) are a major class of flavonoids that contribute to citrus fruit quality. However, the molecular mechanism responsible for PA biosynthesis and accumulation in citrus remains unclear. Here, we identify a PH4-Noemi regulatory complex that regulates proanthocyanidin biosynthesis in citrus. Overexpression of PH4 or Noemi in citrus calli activated the expression of PA biosynthetic genes and significantly increased the PA content. Interestingly, Noemi was also shown to be up-regulated in CsPH4-overexpressing lines compared with wild-type calli. Simultaneously, CsPH4 partially complemented the PA-deficient phenotype of the Arabidopsis tt2 mutant and promoted PA accumulation in the wild-type. Further analysis revealed that CsPH4 interacted with Noemi, and together these proteins synergistically activated the expression of PA biosynthetic genes by directly binding to the MYB-recognizing elements (MRE) of the promoters of these genes. Moreover, CsPH4 could directly bind to the promoter of Noemi and up-regulate the expression of this gene. These findings explain how the CsPH4-Noemi regulatory complex contributes to the activation of PA biosynthetic genes via a positive feedback loop and provide new insights into the molecular mechanisms underlying PA biosynthesis, which can be effectively employed for metabolic engineering to improve citrus fruit quality.
Keywords: Citrus; positive feedback; proanthocyanidin biosynthesis; regulatory complex; transcriptional regulation.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
Similar articles
-
Ectopic Expression of the Coleus R2R3 MYB-Type Proanthocyanidin Regulator Gene SsMYB3 Alters the Flower Color in Transgenic Tobacco.PLoS One. 2015 Oct 8;10(10):e0139392. doi: 10.1371/journal.pone.0139392. eCollection 2015. PLoS One. 2015. PMID: 26448466 Free PMC article.
-
The transcription factor MYB115 contributes to the regulation of proanthocyanidin biosynthesis and enhances fungal resistance in poplar.New Phytol. 2017 Jul;215(1):351-367. doi: 10.1111/nph.14569. Epub 2017 Apr 26. New Phytol. 2017. PMID: 28444797
-
Characterization of two TT2-type MYB transcription factors regulating proanthocyanidin biosynthesis in tetraploid cotton, Gossypium hirsutum.Planta. 2017 Aug;246(2):323-335. doi: 10.1007/s00425-017-2682-z. Epub 2017 Apr 18. Planta. 2017. PMID: 28421329
-
New perspectives on proanthocyanidin biochemistry and molecular regulation.Phytochemistry. 2003 Sep;64(2):367-83. doi: 10.1016/s0031-9422(03)00377-7. Phytochemistry. 2003. PMID: 12943753 Review.
-
Molecular Controls of Proanthocyanidin Synthesis and Structure: Prospects for Genetic Engineering in Crop Plants.J Agric Food Chem. 2018 Sep 26;66(38):9882-9888. doi: 10.1021/acs.jafc.8b02950. Epub 2018 Sep 13. J Agric Food Chem. 2018. PMID: 30139248 Review.
Cited by
-
Weighted gene coexpression correlation network analysis reveals the potential molecular regulatory mechanism of citrate and anthocyanin accumulation between postharvest 'Bingtangcheng' and 'Tarocco' blood orange fruit.BMC Plant Biol. 2023 Jun 2;23(1):296. doi: 10.1186/s12870-023-04309-5. BMC Plant Biol. 2023. PMID: 37268922 Free PMC article.
-
Flavonoids - flowers, fruit, forage and the future.J R Soc N Z. 2022 Feb 28;53(3):304-331. doi: 10.1080/03036758.2022.2034654. eCollection 2023. J R Soc N Z. 2022. PMID: 39439482 Free PMC article. Review.
-
CsTT8 regulates anthocyanin accumulation in blood orange through alternative splicing transcription.Hortic Res. 2023 Sep 19;10(10):uhad190. doi: 10.1093/hr/uhad190. eCollection 2023 Oct. Hortic Res. 2023. PMID: 37927409 Free PMC article.
-
Advances in Biosynthesis and Biological Functions of Proanthocyanidins in Horticultural Plants.Foods. 2020 Nov 30;9(12):1774. doi: 10.3390/foods9121774. Foods. 2020. PMID: 33265960 Free PMC article. Review.
-
Function and transcriptional regulation of CsKCS20 in the elongation of very-long-chain fatty acids and wax biosynthesis in Citrus sinensis flavedo.Hortic Res. 2022 Jan 18;9:uhab027. doi: 10.1093/hr/uhab027. Online ahead of print. Hortic Res. 2022. PMID: 35039844 Free PMC article.
References
-
- Abrahams S, Lee E, Walker AR, Tanner GJ, Larkin PJ, Ashton AR. 2003. The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. The Plant Journal 35, 624–636. - PubMed
-
- Allan AC, Hellens RP, Laing WA. 2008. MYB transcription factors that colour our fruit. Trends in Plant Science 13, 99–102. - PubMed
-
- Amato A, Cavallini E, Walker AR, et al. . 2019. The MYB5-driven MBW complex recruits a WRKY factor to enhance the expression of targets involved in vacuolar hyper-acidification and trafficking in grapevine. The Plant Journal 99, 1220–1241. - PubMed
-
- Bagchi D, Bagchi M, Stohs SJ, Das DK, Ray SD, Kuszynski CA, Joshi SS, Pruess HG. 2000. Free radicals and grape seed proanthocyanidin extract: importance in human health and disease prevention. Toxicology 148, 187–197. - PubMed
-
- Barbehenn RV, Constabel CP. 2011. Tannins in plant-herbivore interactions. Phytochemistry 72, 1551–1565. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
