

Mathematical modelling of telomere length dynamics
- PMID: 31728621
- PMCID: PMC7028829
- DOI: 10.1007/s00285-019-01448-y
Mathematical modelling of telomere length dynamics
Abstract
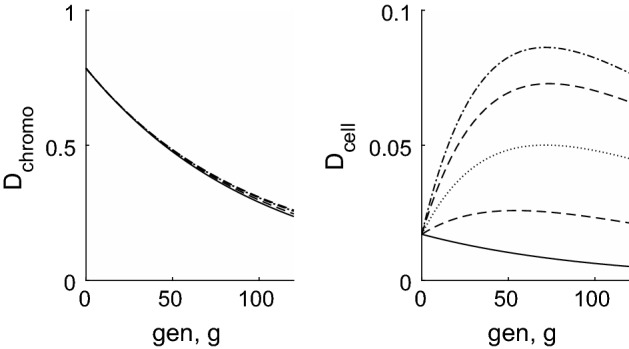
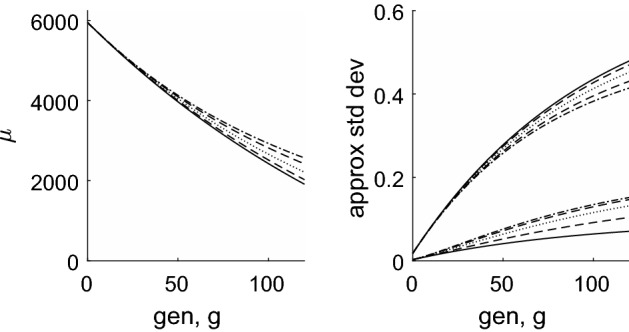
Telomeres are repetitive DNA sequences located at the ends of chromosomes. During cell division, an incomplete copy of each chromosome's DNA is made, causing telomeres to shorten on successive generations. When a threshold length is reached replication ceases and the cell becomes 'senescent'. In this paper, we consider populations of telomeres and, from discrete models, we derive partial differential equations which describe how the distribution of telomere lengths evolves over many generations. We initially consider a population of cells each containing just a single telomere. We use continuum models to compare the effects of various mechanisms of telomere shortening and rates of cell division during normal ageing. For example, the rate (or probability) of cell replication may be fixed or it may decrease as the telomeres shorten. Furthermore, the length of telomere lost on each replication may be constant, or may decrease as the telomeres shorten. Where possible, explicit solutions for the evolution of the distribution of telomere lengths are presented. In other cases, expressions for the mean of the distribution are derived. We extend the models to describe cell populations in which each cell contains a distinct subpopulation of chromosomes. As for the simpler models, constant telomere shortening leads to a linear reduction in telomere length over time, whereas length-dependent shortening results in initially rapid telomere length reduction, slowing at later times. Our analysis also reveals that constant telomere loss leads to a Gaussian (normal) distribution of telomere lengths, whereas length-dependent loss leads to a log-normal distribution. We show that stochastic models, which include a replication probability, also lead to telomere length distributions which are skewed.
Keywords: Aging; End-replication problem; Mathematical; Telomere dynamics.
Figures
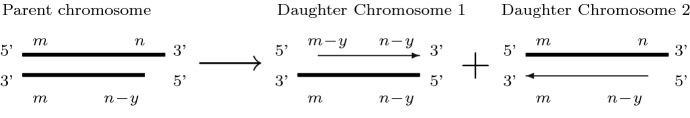
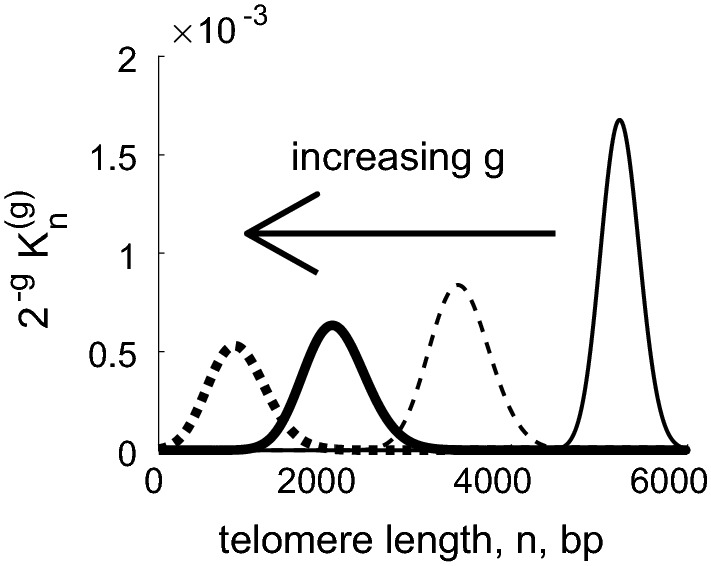
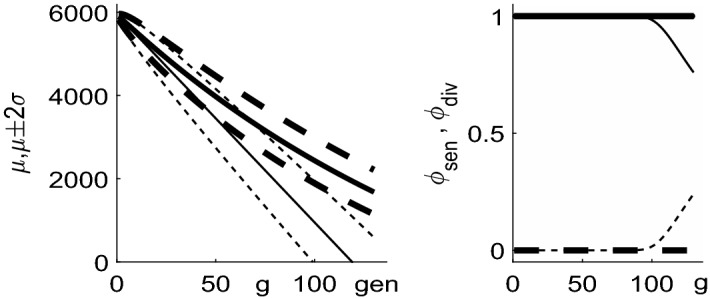
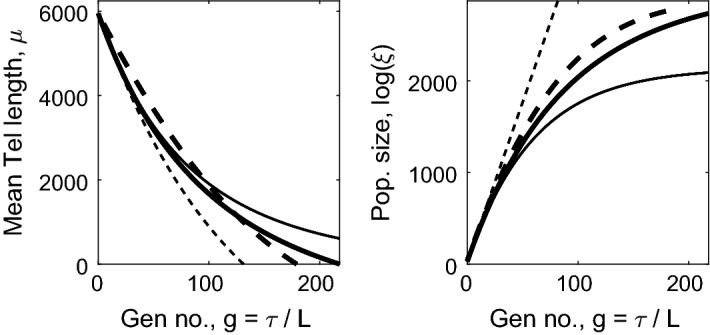
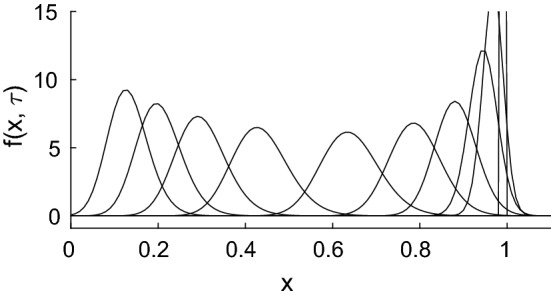
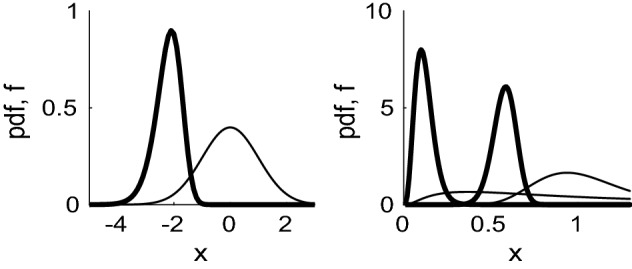
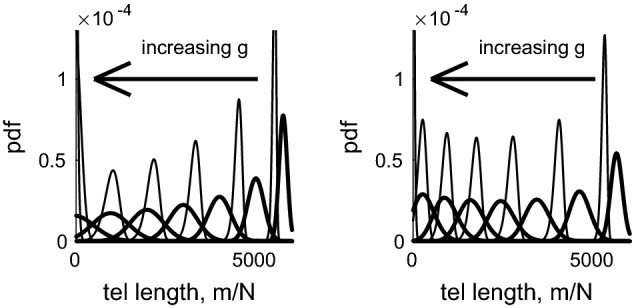
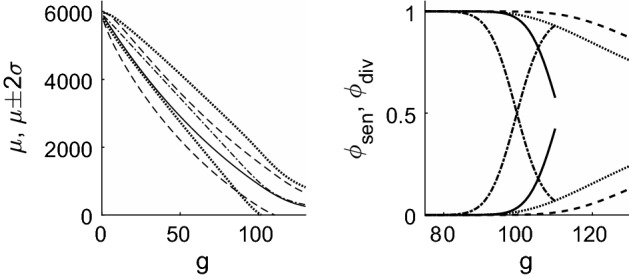
References
-
- Abramowitz M, Stegun I. Handbook of mathematical functions. New York: Dover; 1972.
-
- Bodova K, Kollar R, Tomaska L, Nosek J. Mathematical model of telomere length maintenance in mitochondrial DNA of yeast. Biophys J. 2012;102:281A. doi: 10.1016/j.bpj.2011.11.1555. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
