

Metabolism of Benzalkonium Chlorides by Human Hepatic Cytochromes P450
- PMID: 31730751
- PMCID: PMC7269367
- DOI: 10.1021/acs.chemrestox.9b00293
Metabolism of Benzalkonium Chlorides by Human Hepatic Cytochromes P450
Abstract
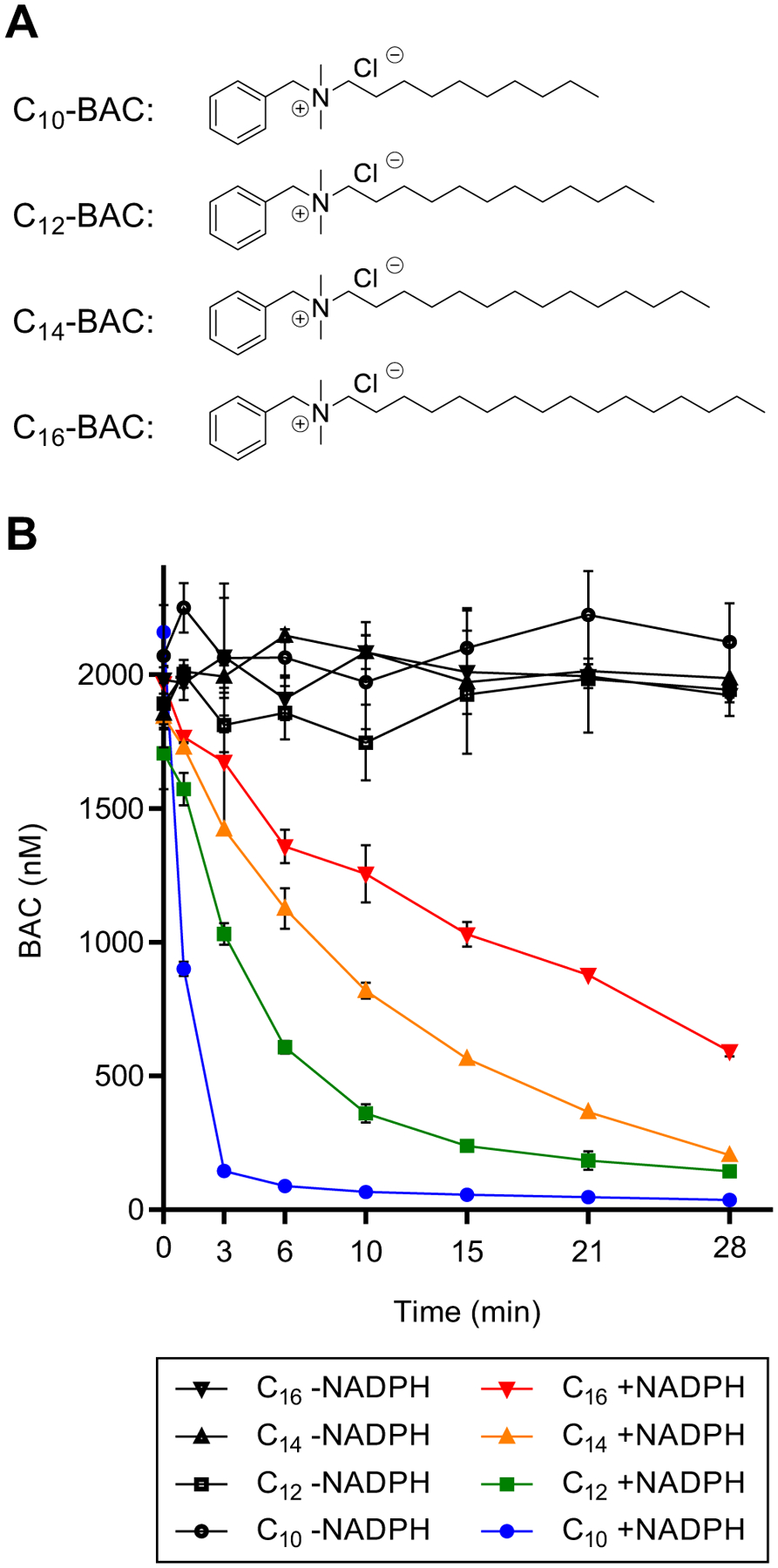
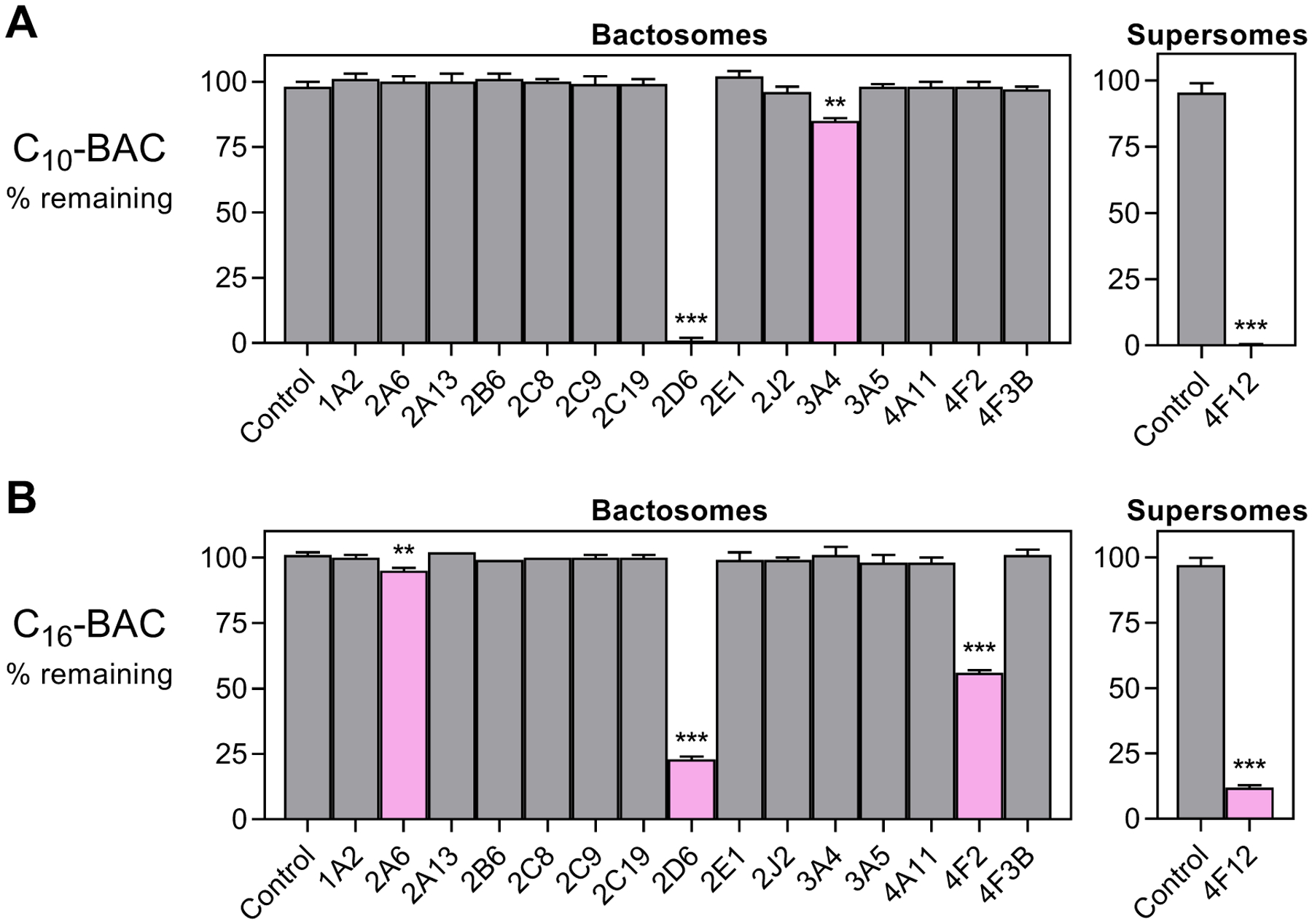
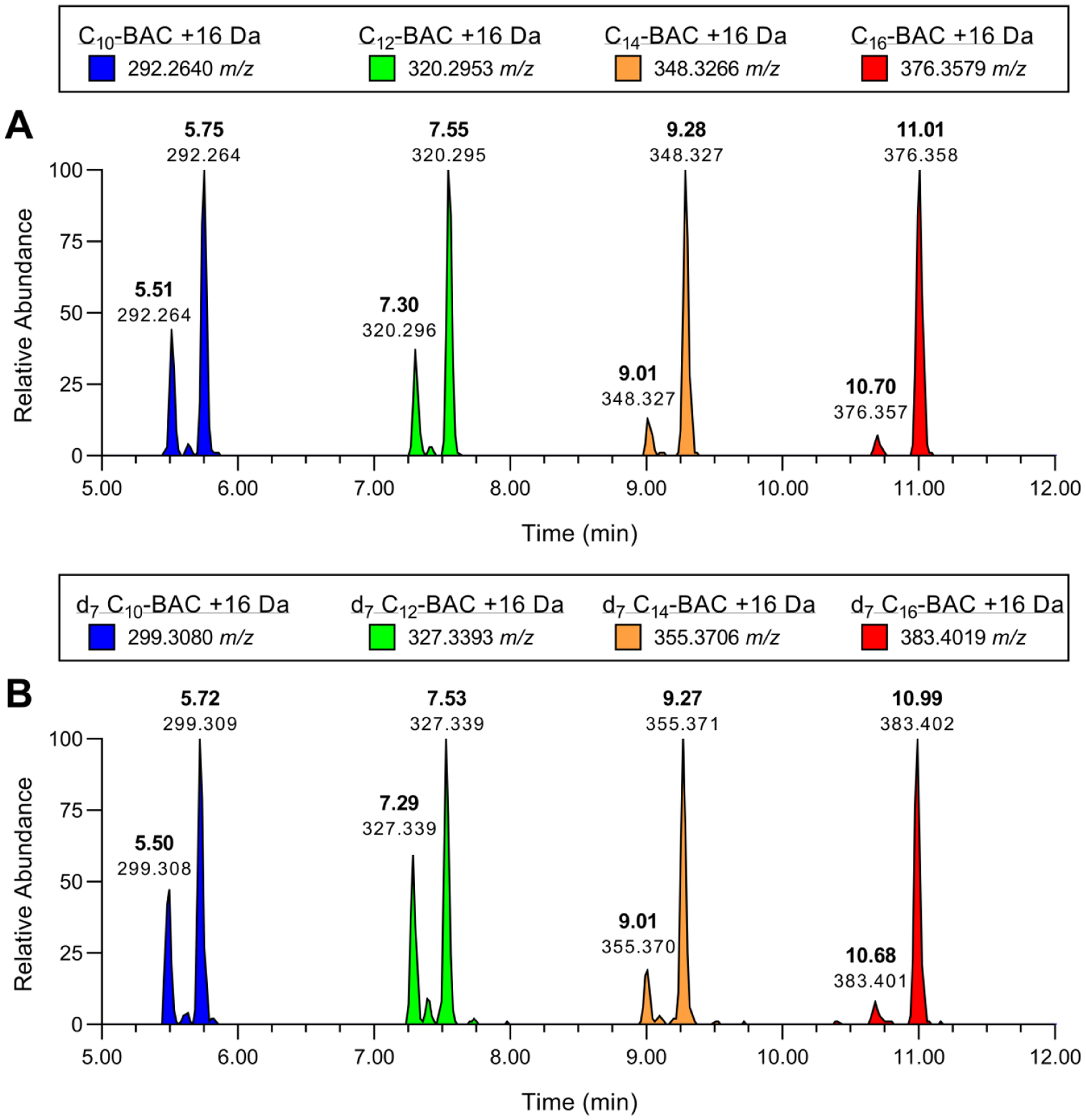
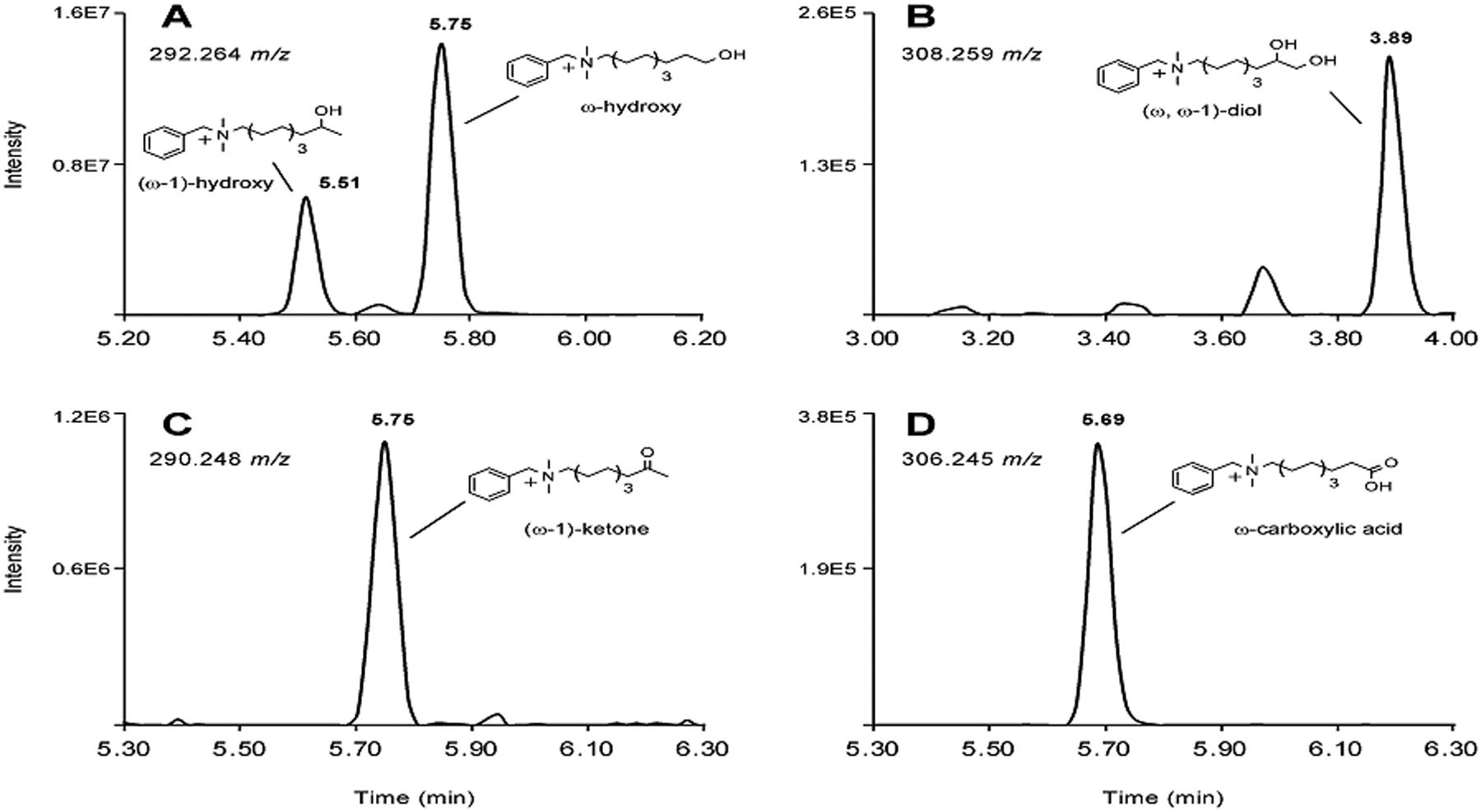
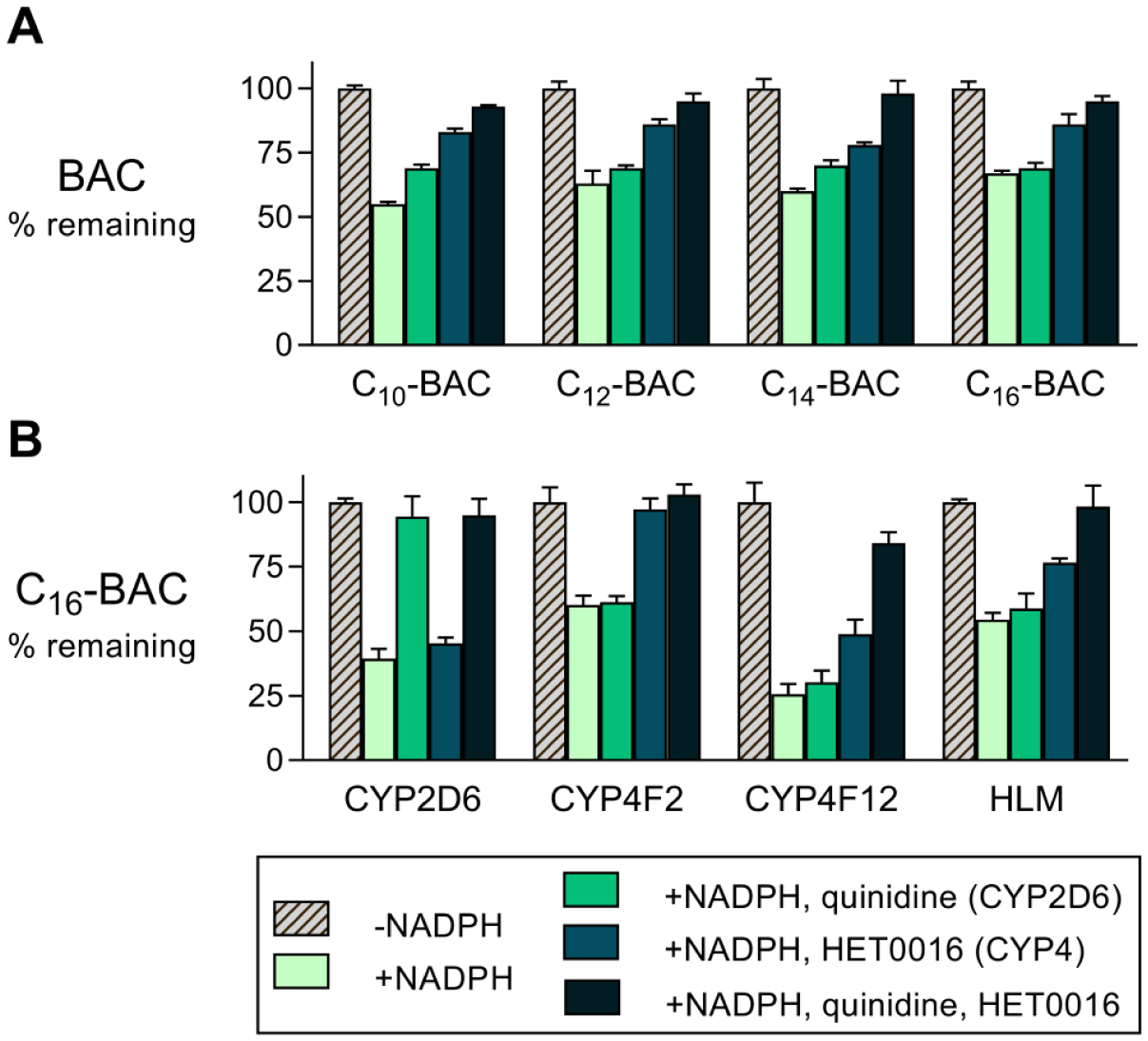
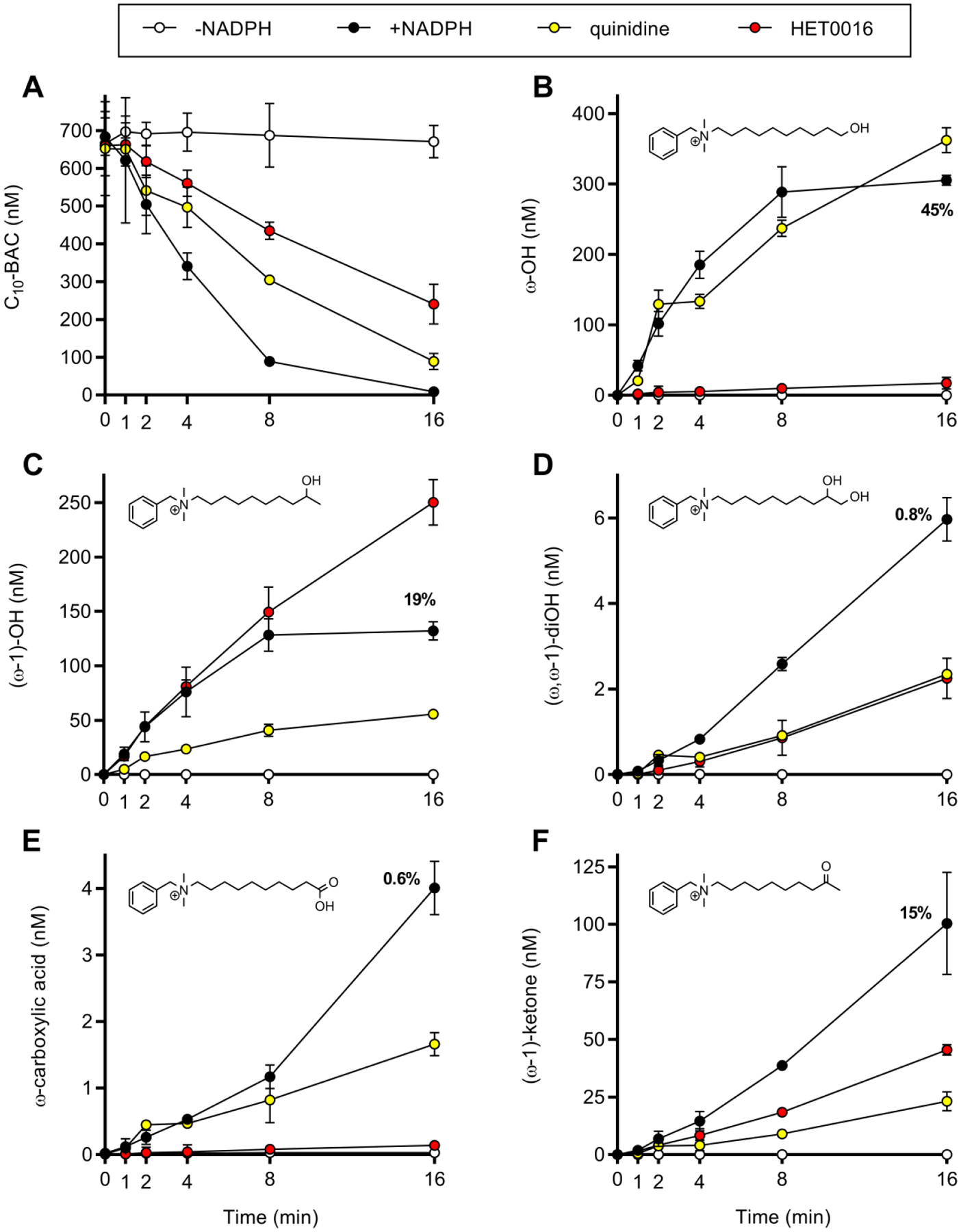
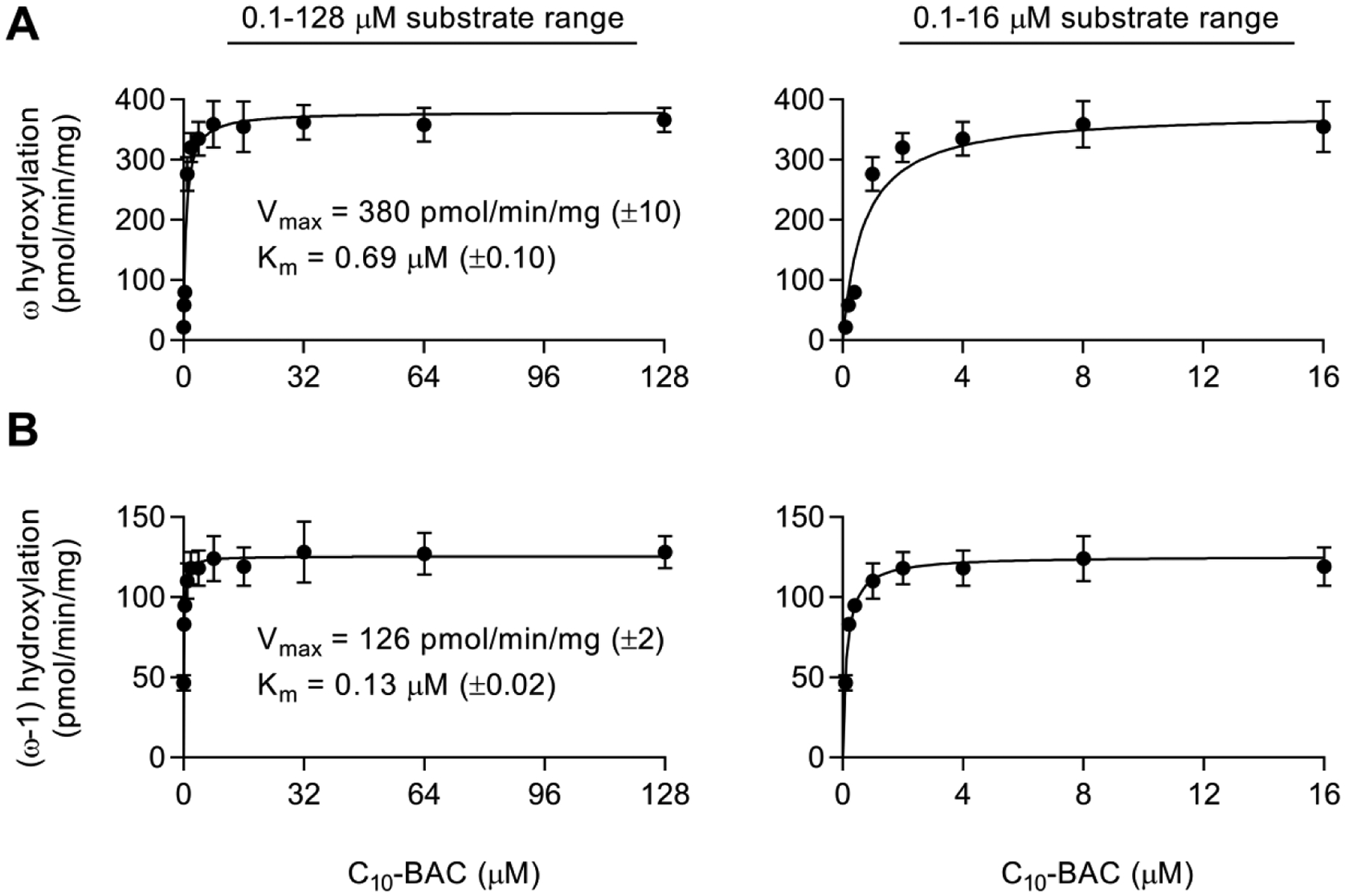
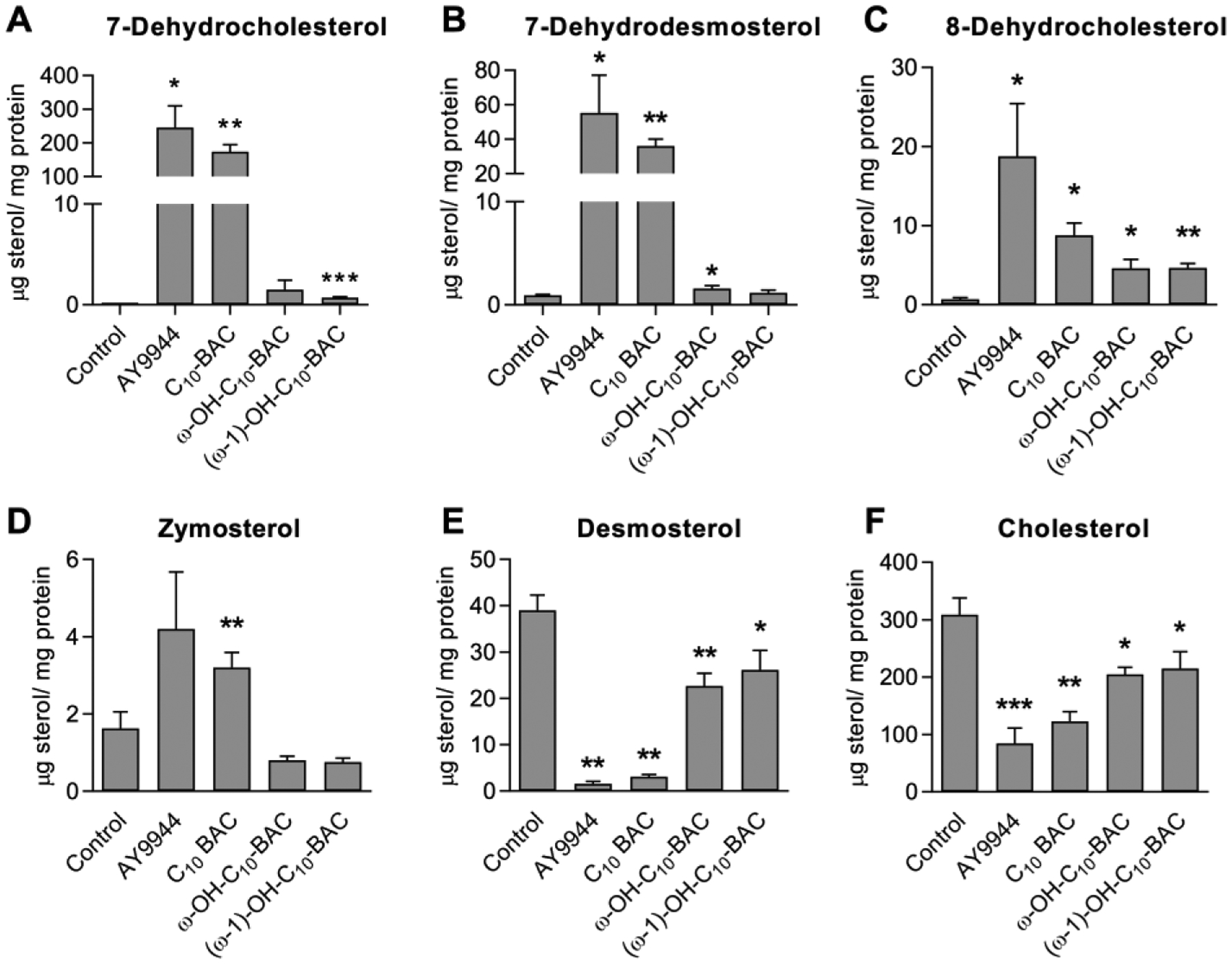
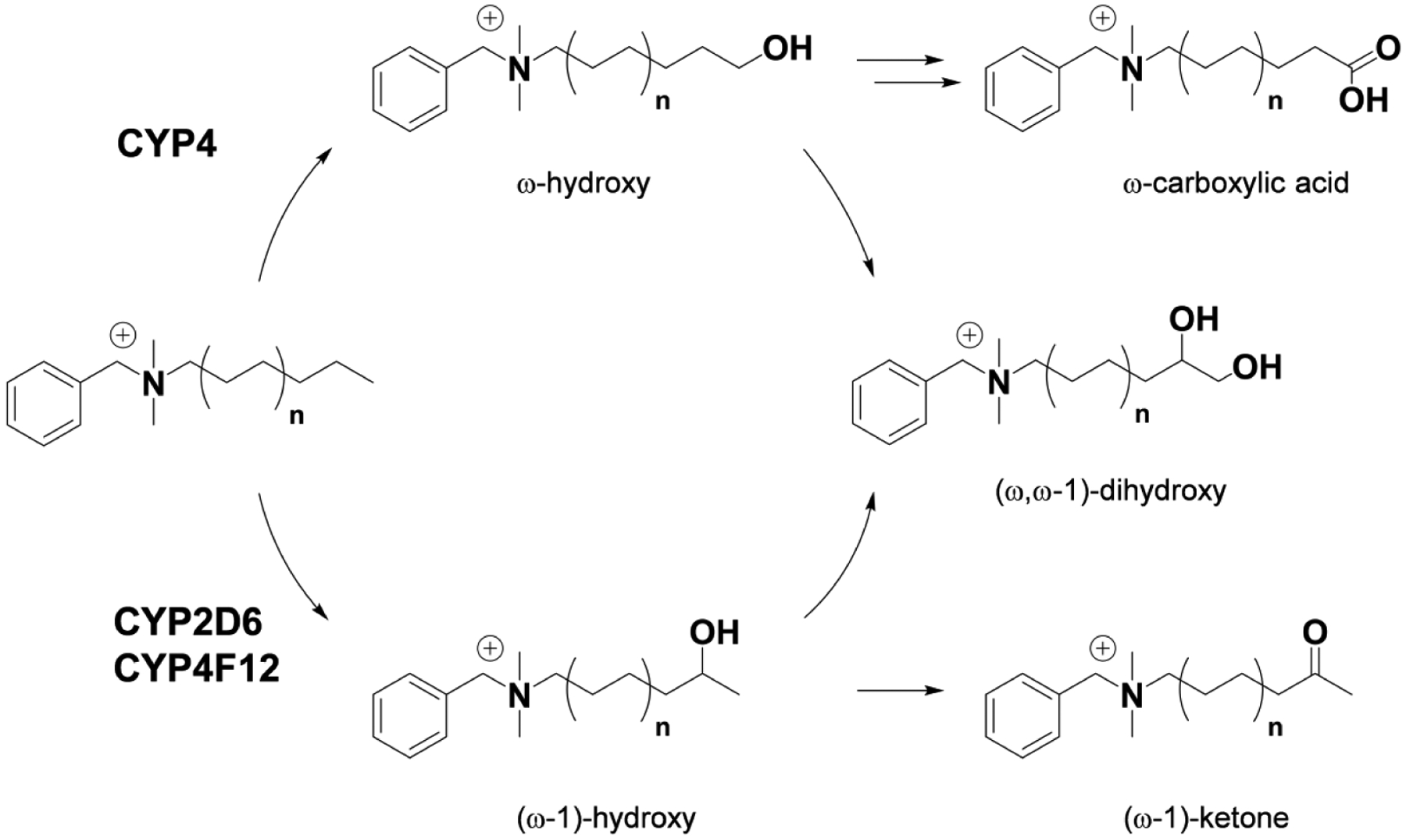
Benzalkonium chlorides (BACs) are widely used as disinfectants in cleaning products, medical products, and the food processing industry. Despite a wide range of reported toxicities, limited studies have been conducted on the metabolism of these compounds in animal models and none in human-derived cells or tissues. In this work, we report on the metabolism of BACs in human liver microsomes (HLM) and by recombinant human hepatic cytochrome P450 (CYP) enzymes. BAC metabolism in HLM was NADPH-dependent and displayed apparent half-lives that increased with BAC alkyl chain length (C10 < C12 < C14 < C16), suggesting enhanced metabolic stability of the more lipophilic, longer chain BACs. Metabolites of d7-benzyl labeled BAC substrates retained all deuteriums and there was no evidence of N-dealkylation. Tandem mass spectrometry fragmentation of BAC metabolites confirmed that oxidation occurs on the alkyl chain region. Major metabolites of C10-BAC were identified as ω-hydroxy-, (ω-1)-hydroxy-, (ω, ω-1)-diol-, (ω-1)-ketone-, and ω-carboxylic acid-C10-BAC by liquid chromatography-mass spectrometry comparison with synthetic standards. In a screen of hepatic CYP isoforms, recombinant CYP2D6, CYP4F2, and CYP4F12 consumed substantial quantities of BAC substrates and produced the major microsomal metabolites. The use of potent pan-CYP4 inhibitor HET0016, the specific CYP2D6 inhibitor quinidine, or both confirmed major contributions of CYP4- and CYP2D6-mediated metabolism in the microsomal disappearance of BACs. Kinetic characterization of C10-BAC metabolite formation in HLM demonstrated robust Michaelis-Menten kinetic parameters for ω-hydroxylation (Vmax = 380 pmol/min/mg, Km = 0.69 μM) and (ω-1)-hydroxylation (Vmax = 126 pmol/min/mg, Km = 0.13 μM) reactions. This work illustrates important roles for CYP4-mediated ω-hydroxylation and CYP2D6/CYP4-mediated (ω-1)-hydroxylation during the hepatic elimination of BACs, an environmental contaminant of emerging concern. Furthermore, we demonstrate that CYP-mediated oxidation of C10-BAC mitigates the potent inhibition of cholesterol biosynthesis exhibited by this short-chain BAC.
Figures
References
-
- Kummerer K, Eitel A, Braun U, Hubner P, Daschner F, Mascart G, Milandri M, Reinthaler F, and Verhoef J (1997) Analysis of benzalkonium chloride in the effluent from European hospitals by solid-phase extraction and high-performance liquid chromatography with post-column ion-pairing and fluorescence detection. J Chromatogr A 774, 281–286. - PubMed
-
- Tezel U, Pierson JA, and Pavlostathis SG (2006) Fate and effect of quaternary ammonium compounds on a mixed methanogenic culture. Water research 40, 3660–3668. - PubMed
-
- Games LM, King JE, and Larson RJ (1982) Fate and Distribution of a Quaternary Ammonium Surfactant, Octadecyltrimethylammonium Chloride (Otac), in Wastewater-Treatment. Environ Sci Technol 16, 483–488.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
