

Genetic Identification of Vagal Sensory Neurons That Control Feeding
- PMID: 31730854
- PMCID: PMC6916730
- DOI: 10.1016/j.cell.2019.10.031
Genetic Identification of Vagal Sensory Neurons That Control Feeding
Abstract
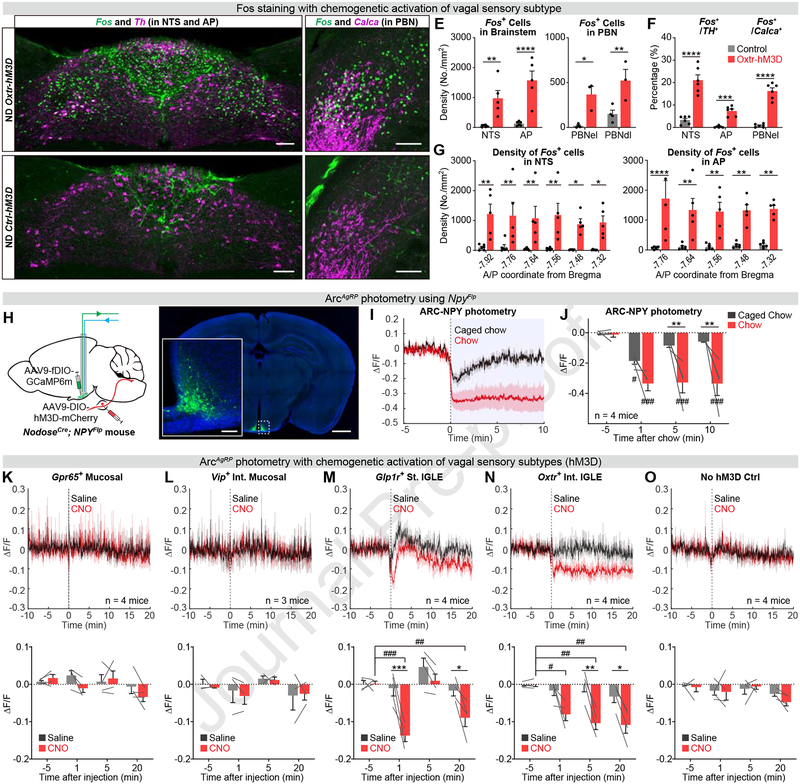
Energy homeostasis requires precise measurement of the quantity and quality of ingested food. The vagus nerve innervates the gut and can detect diverse interoceptive cues, but the identity of the key sensory neurons and corresponding signals that regulate food intake remains unknown. Here, we use an approach for target-specific, single-cell RNA sequencing to generate a map of the vagal cell types that innervate the gastrointestinal tract. We show that unique molecular markers identify vagal neurons with distinct innervation patterns, sensory endings, and function. Surprisingly, we find that food intake is most sensitive to stimulation of mechanoreceptors in the intestine, whereas nutrient-activated mucosal afferents have no effect. Peripheral manipulations combined with central recordings reveal that intestinal mechanoreceptors, but not other cell types, potently and durably inhibit hunger-promoting AgRP neurons in the hypothalamus. These findings identify a key role for intestinal mechanoreceptors in the regulation of feeding.
Keywords: AgRP Neurons; RNA sequencing; chemogenetics; fiber photometry; hypothalamus; optogenetics; satiation; stretch; vagal afferents; vagus nerve.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures






Comment in
-
A good stretch regulates satiety.Nat Rev Neurosci. 2020 Jan;21(1):1. doi: 10.1038/s41583-019-0252-z. Nat Rev Neurosci. 2020. PMID: 31796912 No abstract available.
-
Mechanosensing of food in the gut.Nat Rev Gastroenterol Hepatol. 2020 Feb;17(2):67. doi: 10.1038/s41575-019-0251-6. Nat Rev Gastroenterol Hepatol. 2020. PMID: 31822822 No abstract available.
References
-
- Berthoud H, and Neuhuber WL (2000). Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. Basic Clin. 85, 1–17. - PubMed
-
- Berthoud HR, and Patterson LM (1996). Anatomical relationship between vagal afferent fibers and CCK-immunoreactive entero-endocrine cells in the rat small intestinal mucosa. Acta Anat. (Basel). 156, 123–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
