

Punicalagin Prevents Inflammation in LPS-Induced RAW264.7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway
- PMID: 31731808
- PMCID: PMC6893462
- DOI: 10.3390/nu11112794
Punicalagin Prevents Inflammation in LPS-Induced RAW264.7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway
Abstract
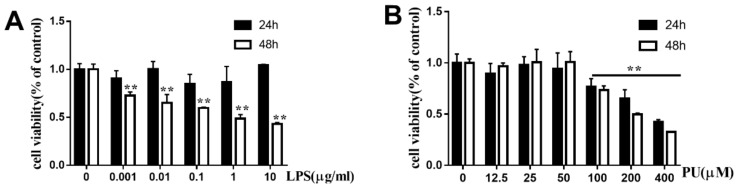
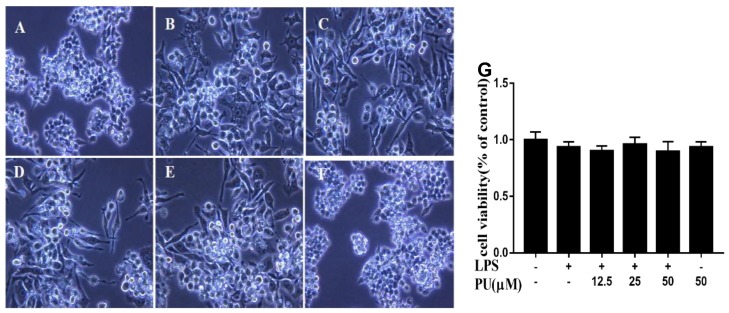
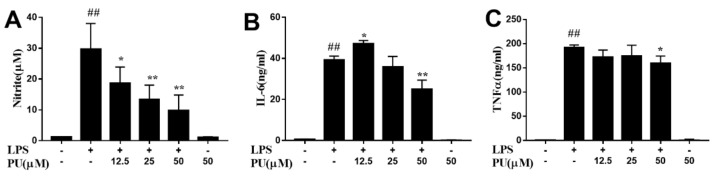
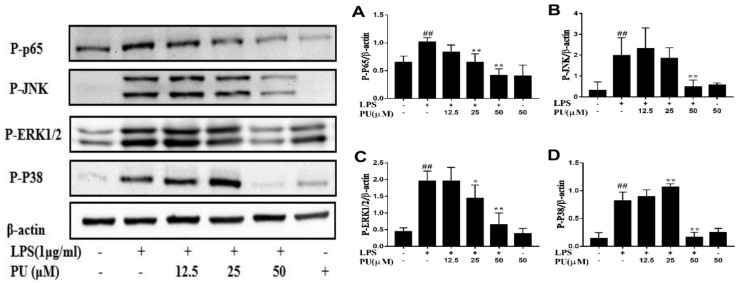
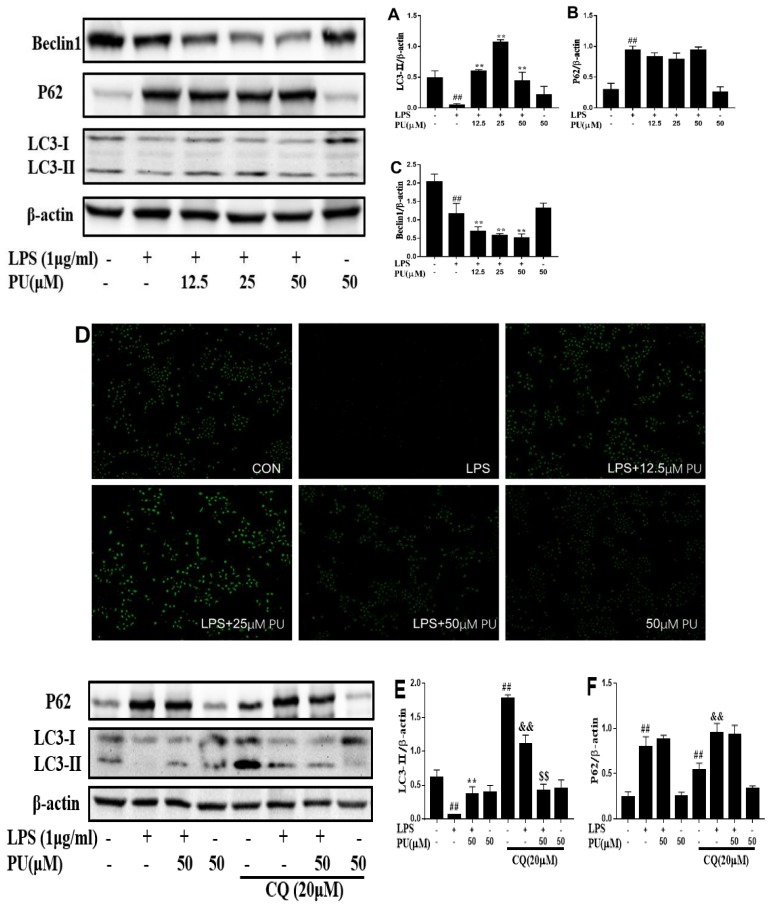
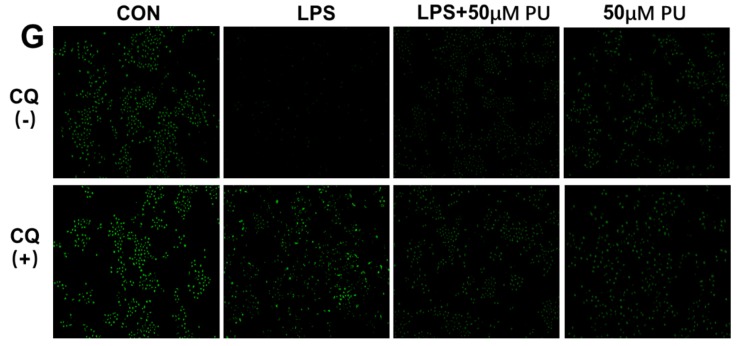
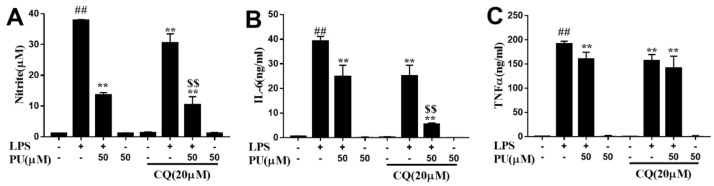
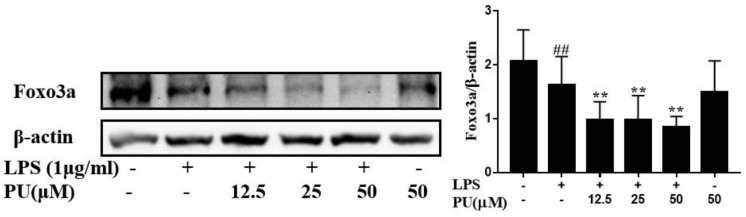
Punicalagin, a hydrolysable tannin of pomegranate juice, exhibits multiple biological effects, including inhibiting production of pro-inflammatory cytokines in macrophages. Autophagy, an intracellular self-digestion process, has been recently shown to regulate inflammatory responses. In this study, we investigated the anti-inflammatory potential of punicalagin in lipopolysaccharide (LPS) induced RAW264.7 macrophages and uncovered the underlying mechanisms. Punicalagin significantly attenuated, in a concentration-dependent manner, LPS-induced release of NO and decreased pro-inflammatory cytokines TNF-α and IL-6 release at the highest concentration. We found that punicalagin inhibited NF-κB and MAPK activation in LPS-induced RAW264.7 macrophages. Western blot analysis revealed that punicalagin pre-treatment enhanced LC3II, p62 expression, and decreased Beclin1 expression in LPS-induced macrophages. MDC assays were used to determine the autophagic process and the results worked in concert with Western blot analysis. In addition, our observations indicated that LPS-induced releases of NO, TNF-α, and IL-6 were attenuated by treatment with autophagy inhibitor chloroquine, suggesting that autophagy inhibition participated in anti-inflammatory effect. We also found that punicalagin downregulated FoxO3a expression, resulting in autophagy inhibition. Overall these results suggested that punicalagin played an important role in the attenuation of LPS-induced inflammatory responses in RAW264.7 macrophages and that the mechanisms involved downregulation of the FoxO3a/autophagy signaling pathway.
Keywords: FoxO3a/autophagy signaling pathway; inflammatory responses; punicalagin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Artesunate Attenuates Pro-Inflammatory Cytokine Release from Macrophages by Inhibiting TLR4-Mediated Autophagic Activation via the TRAF6-Beclin1-PI3KC3 Pathway.Cell Physiol Biochem. 2018;47(2):475-488. doi: 10.1159/000489982. Epub 2018 May 22. Cell Physiol Biochem. 2018. PMID: 29794440
-
O-Methylbulbocapnine and Dicentrine Suppress LPS-Induced Inflammatory Response by Blocking NF-κB and AP-1 Activation through Inhibiting MAPKs and Akt Signaling in RAW264.7 Macrophages.Biol Pharm Bull. 2018;41(8):1219-1227. doi: 10.1248/bpb.b18-00037. Biol Pharm Bull. 2018. PMID: 30068871
-
Pinocembrin attenuates lipopolysaccharide-induced inflammatory responses in Labeo rohita macrophages via the suppression of the NF-κB signalling pathway.Fish Shellfish Immunol. 2016 Sep;56:459-466. doi: 10.1016/j.fsi.2016.07.038. Epub 2016 Aug 2. Fish Shellfish Immunol. 2016. PMID: 27492123
-
Punicalagin in Cancer Prevention-Via Signaling Pathways Targeting.Nutrients. 2021 Aug 9;13(8):2733. doi: 10.3390/nu13082733. Nutrients. 2021. PMID: 34444893 Free PMC article. Review.
-
Physiological and Immune Functions of Punicalagin.Nutrients. 2021 Jun 23;13(7):2150. doi: 10.3390/nu13072150. Nutrients. 2021. PMID: 34201484 Free PMC article. Review.
Cited by
-
Amelioration of colitis progression by ginseng-derived exosome-like nanoparticles through suppression of inflammatory cytokines.J Ginseng Res. 2023 Sep;47(5):627-637. doi: 10.1016/j.jgr.2023.01.004. Epub 2023 Jan 7. J Ginseng Res. 2023. PMID: 37720571 Free PMC article.
-
Regulatory effects of mangiferin on LPS-induced inflammatory responses and intestinal flora imbalance during sepsis.Food Sci Nutr. 2023 Dec 27;12(3):2068-2080. doi: 10.1002/fsn3.3907. eCollection 2024 Mar. Food Sci Nutr. 2023. PMID: 38455195 Free PMC article.
-
Natural products in attenuating renal inflammation via inhibiting the NLRP3 inflammasome in diabetic kidney disease.Front Immunol. 2023 May 5;14:1196016. doi: 10.3389/fimmu.2023.1196016. eCollection 2023. Front Immunol. 2023. PMID: 37215100 Free PMC article. Review.
-
Salvianolic acid A alleviates H2O2-induced endothelial oxidative injury via miR-204-5p.Sci Rep. 2024 May 24;14(1):11931. doi: 10.1038/s41598-024-62556-4. Sci Rep. 2024. PMID: 38789509 Free PMC article.
-
Involvement of the p38 MAPK-NLRC4-Caspase-1 Pathway in Ionizing Radiation-Enhanced Macrophage IL-1β Production.Int J Mol Sci. 2022 Nov 9;23(22):13757. doi: 10.3390/ijms232213757. Int J Mol Sci. 2022. PMID: 36430236 Free PMC article.
References
-
- Bernini R., Barontini M., Cis V., Carastro I., Tofani D., Chiodo R.A., Lupattelli P., Incerpi S. Synthesis and Evaluation of the Antioxidant Activity of Lipophilic Phenethyl Trifluoroacetate Esters by in Vitro ABTS, DPPH and in Cell-Culture DCF Assays. Molecules. 2018;23:208. doi: 10.3390/molecules23010208. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
