

Dendritic Cells Targeting Lactobacillus plantarum Strain NC8 with a Surface-Displayed Single-Chain Variable Fragment of CD11c Induce an Antigen-Specific Protective Cellular Immune Response
- PMID: 31740528
- PMCID: PMC6977127
- DOI: 10.1128/IAI.00759-19
Dendritic Cells Targeting Lactobacillus plantarum Strain NC8 with a Surface-Displayed Single-Chain Variable Fragment of CD11c Induce an Antigen-Specific Protective Cellular Immune Response
Abstract
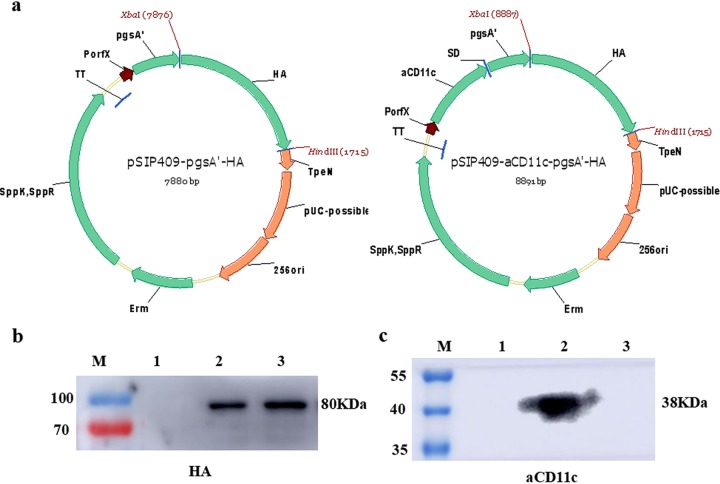
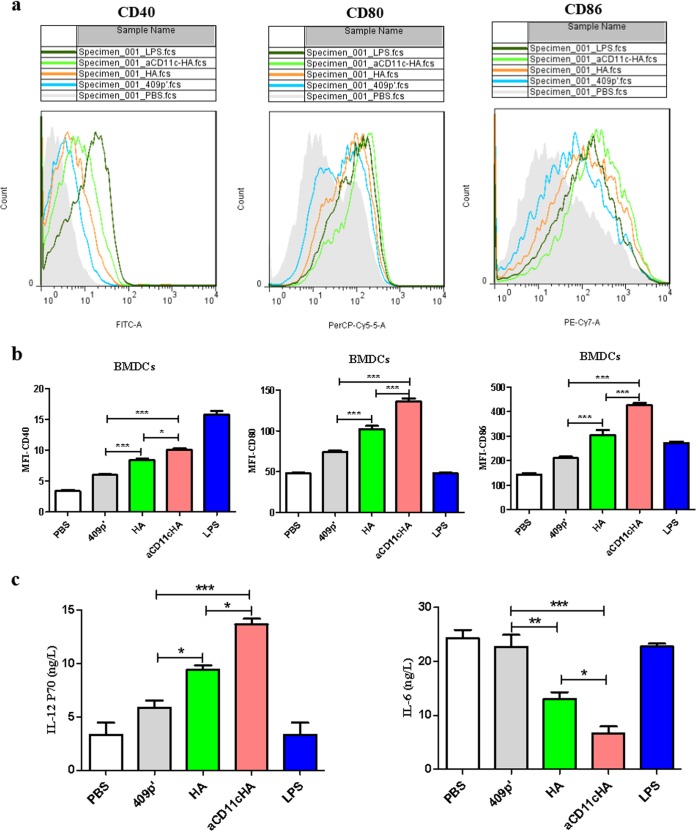
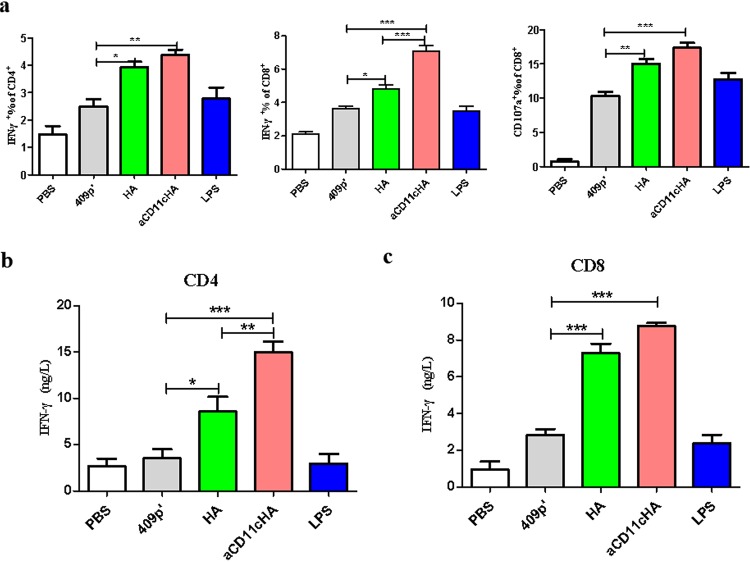
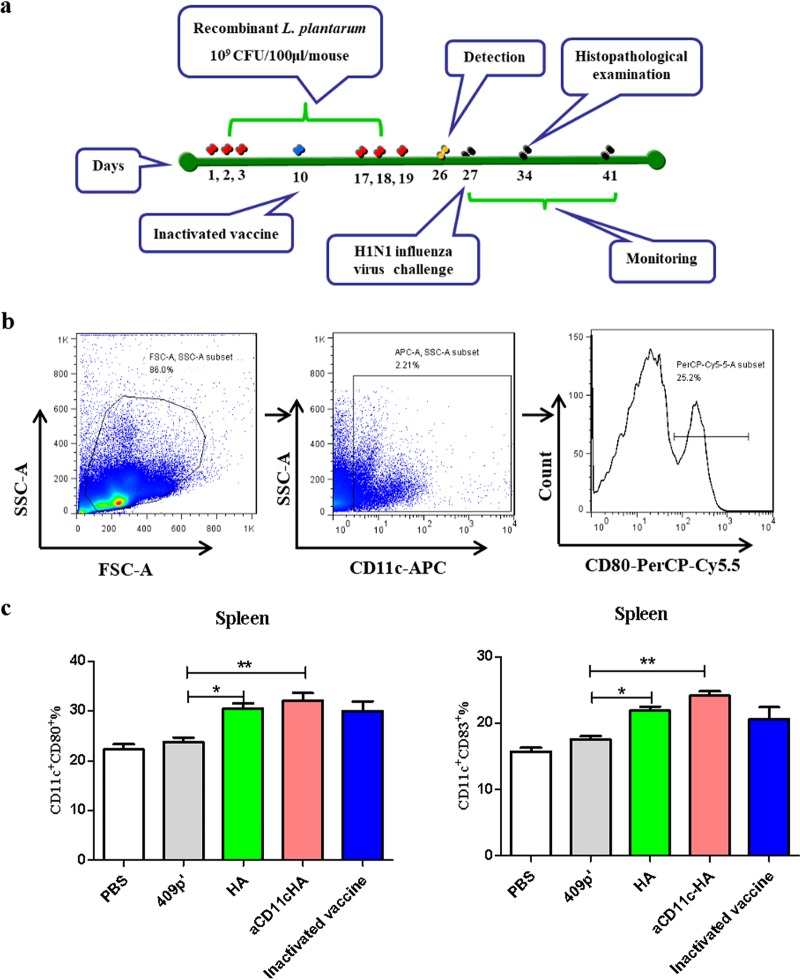
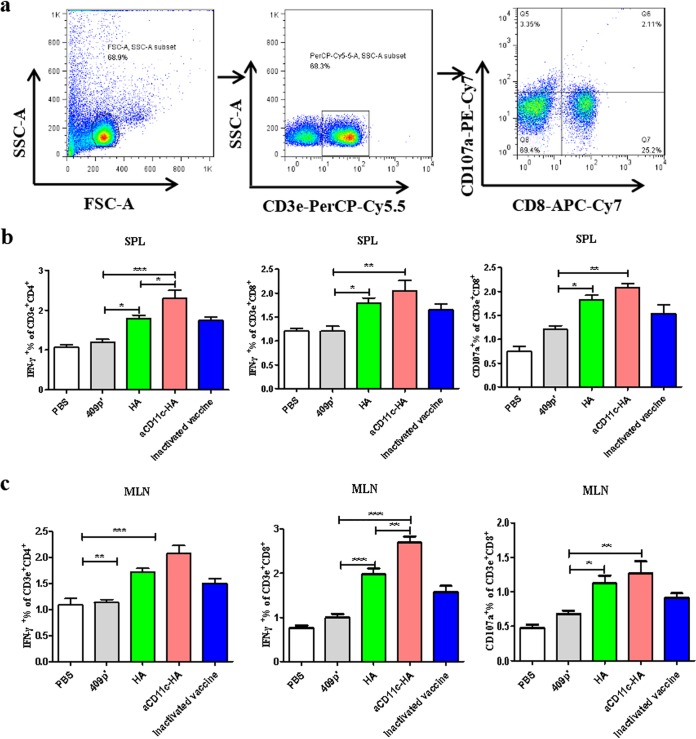
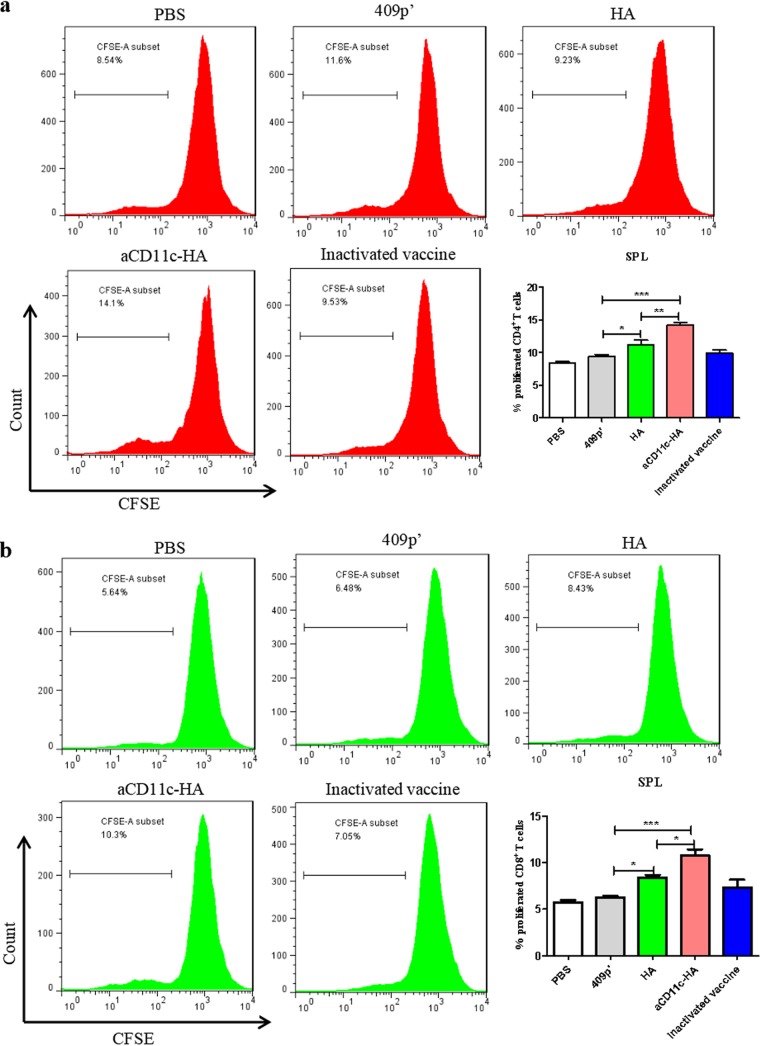
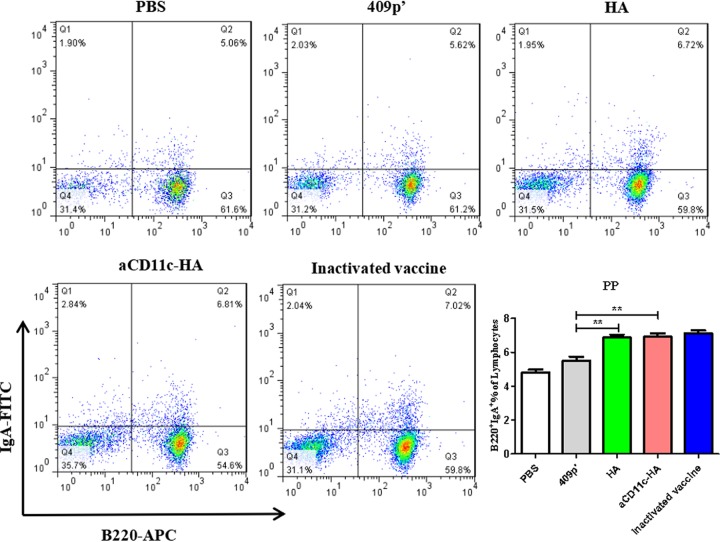
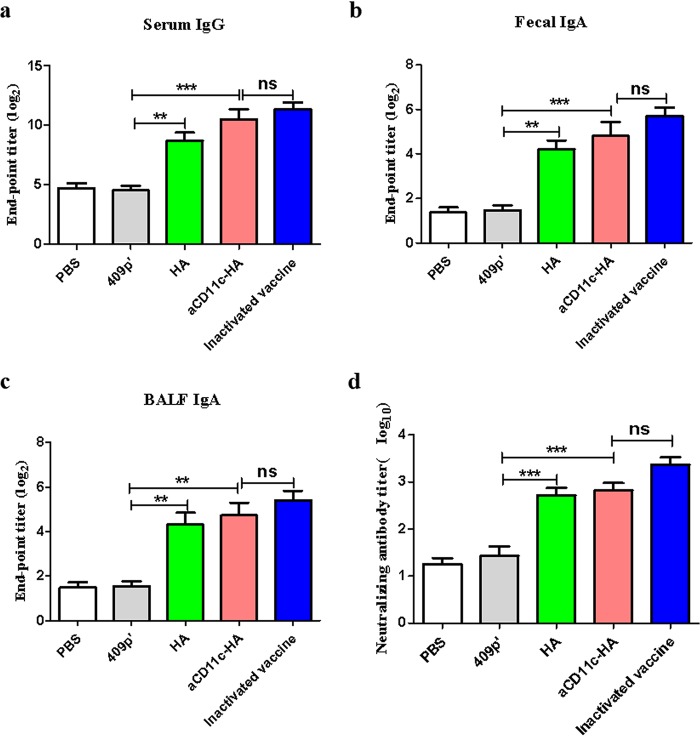
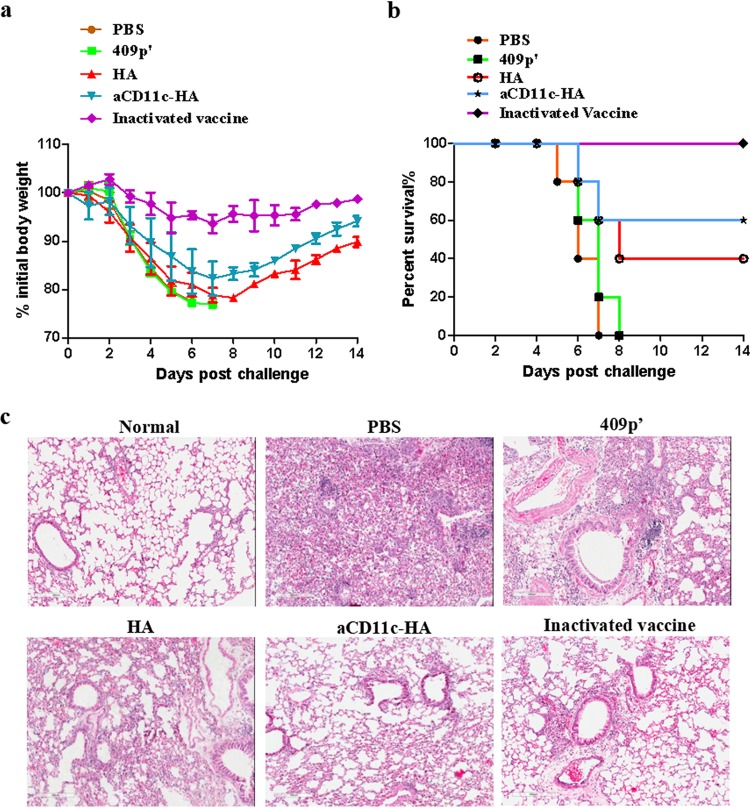
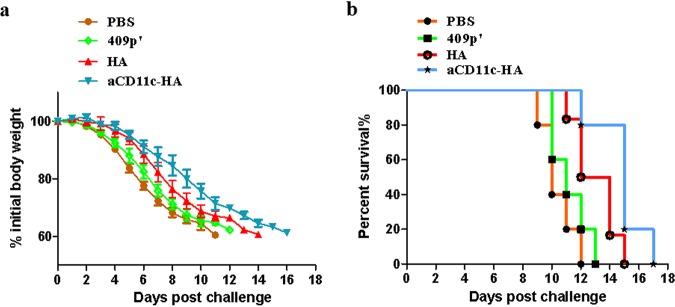
Influenza A virus (H1N1) is an acute, highly contagious respiratory virus. The use of lactic acid bacteria (LAB) to deliver mucosal vaccines against influenza virus infection is a research hot spot. In this study, two recombinant Lactobacillus plantarum strains expressing hemagglutinin (HA) alone or coexpressing aCD11c-HA to target HA protein to dendritic cells (DCs) by fusion to an anti-CD11c single-chain antibody (aCD11c) were constructed. The activation of bone marrow dendritic cells (BMDCs) by recombinant strains and the interaction of activated BMDCs and sorted CD4+ or CD8+ T cells were evaluated through flow cytometry in vitro, and cellular supernatants were assessed by using an enzyme-linked immunosorbent assay kit. The results demonstrated that, compared to the HA strain, the aCD11c-HA strain significantly increased the activation of BMDCs and increased the production of CD4+ gamma interferon-positive (IFN-γ+) T cells, CD8+ IFN-γ+ T cells, and IFN-γ in the cell culture supernatant in vitro Consistent with these results, the aCD11c-HA strain clearly increased the activation and maturation of DCs, the HA-specific responses of CD4+ IFN-γ+ T cells, CD8+ IFN-γ+ T cells, and CD8+ CD107a+ T cells, and the proliferation of T cells in the spleen, finally increasing the levels of specific antibodies and neutralizing antibodies in mice. In addition, the protection of immunized mice was observed after viral infection, as evidenced by improved weight loss, survival, and lung pathology. The adoptive transfer of CD8+ T cells from the aCD11c-HA mice to NOD/Lt-SCID mice resulted in a certain level of protection after influenza virus infection, highlighting the efficacy of the aCD11c targeting strategy.
Keywords: Lactobacillus plantarum; cellular immune response; dendritic cell targeting.
Copyright © 2020 American Society for Microbiology.
Figures
References
-
- Fraser C, Donnelly CA, Cauchemez S, Hanage WP, Van Kerkhove MD, Hollingsworth TD, Griffin J, Baggaley RF, Jenkins HE, Lyons EJ, Jombart T, Hinsley WR, Grassly NC, Balloux F, Ghani AC, Ferguson NM, Rambaut A, Pybus OG, Lopez-Gatell H, Alpuche-Aranda CM, Chapela IB, Zavala EP, Guevara DM, Checchi F, Garcia E, Hugonnet S, Roth C, The WHO Rapid Pandemic Assessment Collaboration . 2009. Pandemic potential of a strain of influenza A (H1N1): early findings. Science 324:1557–1561. doi:10.1126/science.1176062. - DOI - PMC - PubMed
-
- Deng L, Roose K, Job ER, De Rycke R, Van Hamme E, Goncalves A, Parthoens E, Cicchelero L, Sanders N, Fiers W, Saelens X. 2017. Oral delivery of Escherichia coli persistently infected with M2e-displaying bacteriophages partially protects against influenza A virus. J Control Release 264:55–65. doi:10.1016/j.jconrel.2017.08.020. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
